 "
"
Team:Slovenia/ModelingMutualRepressorSwitch
From 2012.igem.org
| Line 518: | Line 518: | ||
</ul> | </ul> | ||
| + | <p> | ||
<!-- figure 1 --> | <!-- figure 1 --> | ||
<table class="invisible" style="width:90%;"> | <table class="invisible" style="width:90%;"> | ||
| Line 536: | Line 537: | ||
</table> | </table> | ||
<!-- end table--> | <!-- end table--> | ||
| + | </p> | ||
</p> | </p> | ||
Revision as of 20:58, 25 September 2012
Modeling - mutual repressor switch
- Deterministic model
- Stochastic model
- C#Sim model
Deterministic model of the mutual repressor switch
|
Deterministic analysis demonstrated that while theoretical conditions under which the mutual repressor switch would exhibit bistability exist, they are unlikely to occur in a realistic experimental setting. This was because:
Increase in cooperativity lead to more robust behavior – higher expression levels of stable states were reached and higher – but still low - tolerance to leaky expression was observed. Experimental results confirmed that the mutual repressor switch did not exhibit bistability. |
The model
We can describe the relations for the mutual repressor switch by the following equations. Fractional occupancies of promoters are:
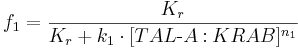
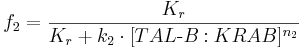
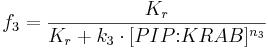
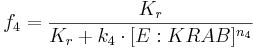
- f1, f2, f3 and f4 are probabilities of promoters 1 (construct 1), 2 (construct 2), 3 (construct 3) and 4 (construct 4), respectively, being in an active state, resulting in gene expression;
- [TAL-A:KRAB], [TAL-B:KRAB], [PIP:KRAB] and [E:KRAB] are protein concentrations at a given time;
- k1, k2, k3 and k4 are association constants;
- n1, n2, n3 and n4 are exponents representing the degree of functional cooperativity;
- Kr is the amount of repressor required for 50% repression of constitutive promoter (equal to 1 in our simulations);
ODEs representing protein production are described by a set of equations:
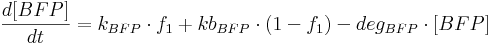
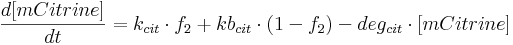
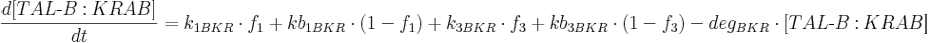
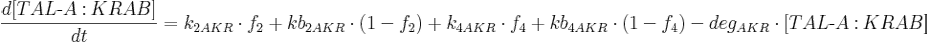
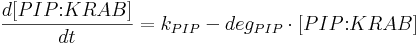
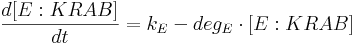
- [BFP], [mCitrine], [TAL-A:KRAB], [TAL-B:KRAB], [PIP:KRAB] and [E:KRAB] are protein concentrations;
- kBFP is BFP production rate from construct 1 (i.e. production rate when construct 1 promoter is active);
- kbBFP is basal BFP production rate from construct 1 (i.e. production rate when construct 1 promoter is inactive);
- degBFP is BFP degradation rate;
- kcit is mCitrine production rate from construct 2 (i.e. production rate when construct 2 promoter is active);
- kbcit is basal mCitrine production rate from construct 2 (i.e. production rate when construct 2 promoter in inactive);
- degcit is mCitrine degradation rate;
- k2AKR is TAL-A:KRAB production rate from construct 2;
- kb2AKR is basal TAL-A:KRAB production rate from construct 2;
- k4AKR is TAL-A:KRAB production rate from construct 4;
- kb4AKR is basal TAL-A:KRAB production rate from construct 4;
- degAKR is TAL-A:KRAB degradation rate;
- k1BKR is TAL-B:KRAB production rate from construct 1;
- kb1BKR Is basal TAL-B:KRAB production rate from construct 1;
- k3BKR is TAL-B:KRAB production rate from construct 3;
- kb3BKR is basal TAL-B:KRAB production rate from construct 3;
- degBKR is TAL-B:KRAB degradation rate;
- kPIP is PIP:KRAB production rate;
- degPIP is PIP:KRAB degradation rate;
- kE is E:KRAB production rate;
- degE is E:KRAB degradation rate.
See model derivation for details.
Simulation results
Simulation results are shown as reporter (BFP, mCitrine) concentrations – indicating one of the two states - as a function of time. No specific units were used, hence no absolute interpretation of the results' values in terms of units is in place. Switching between states was achieved using two signals (signal 1 and signal 2), modeled as a unit step present for a certain duration of time. Each signal was used to induce a different stable state. The presence of signal 1 caused PIP:KRAB to dissociate from promoter 3 binding site and activated transcription. The presence of signal 2 caused E:KRAB to dissociate from promoter 4 binding site and activated transcription.
Initial species' concentrations were 0 for all simulations.
Detailed parameter values for each simulation can be found in MATLAB files here.
Simulation set 1 - switching between stable states
The purpose of this simulation was to show that conditions exist in deterministic setting where the switch can assume each of the two stable states. Functional cooperativity of each transcription factor (TF) was initialy taken as 2.5. Product production:degradation ratio was equal to 100 (i.e. maximal possible expression level was 100). Ideal circumstances with zero leaky expression were assumed. Figure 1 shows the result of this simulation, demonstrating bistability. The following state-switching scenario was used:
- at time = 0, signal 1 was introduced (signal 1 = 1) to induce the first stable state, depicted by a high level of BFP;
- at time = 100, signal 1 was removed from the system (signal 1 = 0); high-BFP state persisted even after that, meaning a stable state was achieved;
- at time = 400, signal 2 was introduced (signal 2 = 1) to induce the second, high-mCitrine stable state;
- at time = 500, signal 2 was removed from the system (signal 2 = 0); high-mCitrine state persisted, meaning a stable state was achieved;
- at time = 800, signal 1 was re-introduced (signal 1 = 1) to show that a transition to high-BFP state could follow a high-mCitrine state;
- at time = 900, signal 1 was removed (signal 1 = 0); high-BFP stable state persisted.
 |
| Figure 1. Mutual repressor switch transitioning between stable states. Reporter (BFP, mCitrine) concentrations are depicted as a function of time. Here, stable states were observed under ideal conditions with no leaky expression and functional cooperativity (exponent values) equal to 2.5. |
Next: Stochastic model of the mutual repressor switch >>