 "
"
Team:British Columbia/Consortia
From 2012.igem.org
| (31 intermediate revisions not shown) | |||
| Line 23: | Line 23: | ||
<br/><br/> | <br/><br/> | ||
The following sections detail the mathematical foundation of the model, the experimental measurement of the necessary constants, the MATLAB implementation and some preliminary results.<br/><br/> | The following sections detail the mathematical foundation of the model, the experimental measurement of the necessary constants, the MATLAB implementation and some preliminary results.<br/><br/> | ||
| - | |||
<font face=arial narrow size=4><b>Equations and Constants</b></font></br><font face=arial narrow></br> | <font face=arial narrow size=4><b>Equations and Constants</b></font></br><font face=arial narrow></br> | ||
| - | |||
| - | |||
We proposed the following fundamental equations for our consortium model. In this model, each auxotrophic strain is denoted with a superscript minus (i.e. <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> are denoted trpA<sup>-</sup>, metA<sup>-</sup>, and tryA<sup>-</sup>, respectively); [Trp], [Met], and [Tyr] represent the concentrations of tryptophan, methionine, and tyrosine; ODtrpA<sup>-</sup>, ODmetA<sup>-</sup>, and ODtyrA<sup>-</sup> represent the optical densities of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains; a1, a2, and a3 are the consumption constants with respect to changes in ODtrpA<sup>-</sup>, ODmetA<sup>-</sup>, and ODtyrA<sup>-</sup>, respectively; µtrpA<sup>-</sup>, µmetA<sup>-</sup>, µtyrA<sup>-</sup> are the specific growth rates of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains; µMaxtrpA<sup>-</sup>, µMaxmetA<sup>-</sup>, µMaxtyrA<sup>-</sup> are the maximum growth rates of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains; and KstrpA<sup>-</sup>, KsmetA<sup>-</sup>, KStyrA<sup>-</sup> are the <i>half-velocity constants</i> of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains. The terms and constants of this equation will be discussed in further detail later.</br></br> | We proposed the following fundamental equations for our consortium model. In this model, each auxotrophic strain is denoted with a superscript minus (i.e. <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> are denoted trpA<sup>-</sup>, metA<sup>-</sup>, and tryA<sup>-</sup>, respectively); [Trp], [Met], and [Tyr] represent the concentrations of tryptophan, methionine, and tyrosine; ODtrpA<sup>-</sup>, ODmetA<sup>-</sup>, and ODtyrA<sup>-</sup> represent the optical densities of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains; a1, a2, and a3 are the consumption constants with respect to changes in ODtrpA<sup>-</sup>, ODmetA<sup>-</sup>, and ODtyrA<sup>-</sup>, respectively; µtrpA<sup>-</sup>, µmetA<sup>-</sup>, µtyrA<sup>-</sup> are the specific growth rates of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains; µMaxtrpA<sup>-</sup>, µMaxmetA<sup>-</sup>, µMaxtyrA<sup>-</sup> are the maximum growth rates of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains; and KstrpA<sup>-</sup>, KsmetA<sup>-</sup>, KStyrA<sup>-</sup> are the <i>half-velocity constants</i> of the <i>ΔtrpA</i>, <i>ΔmetA</i>, and <i>ΔtyrA</i> strains. The terms and constants of this equation will be discussed in further detail later.</br></br> | ||
| Line 39: | Line 36: | ||
Applying the Monod kinetic model, we sought out to measure three variables: the concentration of tryptophan, methionine, and tyrosine in the media available for each specific auxotroph. We were also interested in how these concentrations vary with respect to time. </br></br> | Applying the Monod kinetic model, we sought out to measure three variables: the concentration of tryptophan, methionine, and tyrosine in the media available for each specific auxotroph. We were also interested in how these concentrations vary with respect to time. </br></br> | ||
| - | <p align=center><img src="https://static.igem.org/mediawiki/2012/3/3d/GrowthRatesAA_university_of_british_columbia.png"></br><b>Figure 1: Monoculture Growth Rates at various Limiting Amino Acid Concentrations. These data are fit to the Monod equation to determine the constants listed below.</br></br> | + | <p align=center><img src="https://static.igem.org/mediawiki/2012/3/3d/GrowthRatesAA_university_of_british_columbia.png"></br><b>Figure 1: Monoculture Growth Rates at various Limiting Amino Acid Concentrations. |
| - | <img src="https://static.igem.org/mediawiki/2012/0/05/GrowthRatesAA_LOG_university_of_british_columbia.png"></br> | + | |
| + | <p align=center></br></br>These data are fit to the Monod equation to determine the constants listed below.</br></br> | ||
| + | <p align=center><img src="https://static.igem.org/mediawiki/2012/0/05/GrowthRatesAA_LOG_university_of_british_columbia.png"></br> | ||
Figure 2: Log Scale of Monoculture Growth Rates at various Limiting Amino Acid Concentrations.</b></br></br> | Figure 2: Log Scale of Monoculture Growth Rates at various Limiting Amino Acid Concentrations.</b></br></br> | ||
| - | + | </br></br> | |
| + | <p align=left>Note: we used lower amino acid concentrations as well, however we found out that when concentration is below 1E<sup>-7</sup> M, we observed no significant overnight growth of the cells. For a detailed protocol, please visit <a href="https://2012.igem.org/Team:British_Columbia/Protocols/Monod">here</a>.</br></br> | ||
Based on the fits presented in Figures 1 and 2, the key constants, such as maximum growth rates, and half-velocity constants, for each auxotroph were obtained [1]:</br></br><p align=center> | Based on the fits presented in Figures 1 and 2, the key constants, such as maximum growth rates, and half-velocity constants, for each auxotroph were obtained [1]:</br></br><p align=center> | ||
<img src="https://static.igem.org/mediawiki/2012/b/b8/British_Columbia_2012_constants_from_GrowthRates.png" width=400></p> | <img src="https://static.igem.org/mediawiki/2012/b/b8/British_Columbia_2012_constants_from_GrowthRates.png" width=400></p> | ||
| - | As discussed in the beginning of this section, we assumed that the amino acid consumption is solely dependent on the cell growth rate. In order to find out the consumption rate constants, (a<sub>1</sub>, a<sub>2</sub>, and a<sub>3</sub>), we need to find out a way to measure the amino acid concentrations in the media with respect to the change of its OD for each auxotroph. To do this, we need to find a way to measure media amino acid concentrations. With no good access to HPLC, we noticed in previous experiments that each of the knockout monocultures will reach to a certain final OD at a different known concentration of amino acid. In addition, recently published paper: ''A Programmable <i>Escherichia coli</i> Consortium via Tunable Symbiosis'', Kerner A et. al (2012) confirmed that there is indeed a relationship between the final auxotroph ODs with the environmental amino acid concentrations [2]. We thus generated a calibration curve to relate the limiting amino acid concentrations to their final OD readings. We based this on our own wet lab data shown in the figures below: </br></br> | + | <p align=left>As discussed in the beginning of this section, we assumed that the amino acid consumption is solely dependent on the cell growth rate. In order to find out the consumption rate constants, (a<sub>1</sub>, a<sub>2</sub>, and a<sub>3</sub>), we need to find out a way to measure the amino acid concentrations in the media with respect to the change of its OD for each auxotroph. To do this, we need to find a way to measure media amino acid concentrations. With no good access to HPLC, we noticed in previous experiments that each of the knockout monocultures will reach to a certain final OD at a different known concentration of amino acid. In addition, recently published paper: ''A Programmable <i>Escherichia coli</i> Consortium via Tunable Symbiosis'', Kerner A et. al (2012) confirmed that there is indeed a relationship between the final auxotroph ODs with the environmental amino acid concentrations [2]. We thus generated a calibration curve to relate the limiting amino acid concentrations to their final OD readings. We based this on our own wet lab data shown in the figures below: </br></br> |
| - | <p align=center><img src="https://static.igem.org/mediawiki/2012/b/be/ | + | <p align=center><img src="https://static.igem.org/mediawiki/2012/b/be/ODcalibrationMet_university_of_british_columbia.png"width=600> |
| - | </br><b>Figure 3:Calibration curves used to convert final OD600 to initial amino acid concentration. Standard solutions of known amino acid concentrations were innoculated with the respective auxotroph and grown for 20 hours at 37oC shaken at 200RPM, after which final OD600 were measured. Data were fit to quartic polynomials to inter- polate OD600 readings. Error bars represent standard deviation (n=3). </b></br></br> | + | <p align=left></br><b>Figure 3: Calibration curves used to convert final OD600 to initial amino acid concentration. Standard solutions of known amino acid concentrations were innoculated with the respective auxotroph and grown for 20 hours at 37oC shaken at 200RPM, after which final OD600 were measured. Data were fit to quartic polynomials to inter- polate OD600 readings. Error bars represent standard deviation (n=3).</b></br></br>. |
| + | <p align=left>We then sought out to find out how the amino acid in the environment is consumed with respect to cell growth. </br></br> | ||
| - | + | <p align=left>The three auxotrophs were grown in 50 ml flasks with known initial Tryptophan, Methionine, and Tyrosine concentrations (5E<sup>-5</sup> M), respectively, and 5ml of cell free supernatants at various ODs was isolated. These supernatants served as new 96-well plate media for each knock out cultures with newly added M9-Glucose shots (2% glucose in the well) to ensure the amino acids of interest remained as the limiting substrate for all knock outs. A detailed protocol can be found <a href="https://2012.igem.org/Team:British_Columbia/Protocols/AArate">here</a>. </br></br> | |
| - | < | + | |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | The three auxotrophs were grown in 50 ml flasks with known initial Tryptophan, Methionine, and Tyrosine concentrations (5E<sup>-5</sup> M), respectively, and 5ml of cell free supernatants at various ODs was isolated. These supernatants served as new 96-well plate media for each knock out cultures with newly added M9-Glucose shots (2% glucose in the well) to ensure the amino acids of interest remained as the limiting substrate for all knock outs. A detailed protocol can be found <a href="https://2012.igem.org/Team:British_Columbia/Protocols/AArate">here</a>. </br></br> | + | |
The cells were grown in a 96-well plate for 15 hours at 37 <sup>o</sup>C, and growth curve data was obtained using a plate reader (most of the monocultures reached stationary phase at this point). </br></br> | The cells were grown in a 96-well plate for 15 hours at 37 <sup>o</sup>C, and growth curve data was obtained using a plate reader (most of the monocultures reached stationary phase at this point). </br></br> | ||
| - | Based on the calibration curves (Figure 3 | + | Based on the calibration curves (Figure 3), the amino acid concentrations in the wells were calculated from the final OD readings, and when corrected with a dilution factor of 5/4 (as we added in the 5X, it was 1/5 of the total well media volume, as described <a href="https://2012.igem.org/Team:British_Columbia/Protocols/AArate">here</a>), we obtained the resultant amino acid concentrations in the supernatant at various auxotroph monoculture densities. Again, with the assumption that the only change in the amino acid concentration in the media is due to cell growth, and as they are specific knock outs, they were not able to release any amino acid into the environment, each auxotrophs' consumption constants with respect to cell growth rate (a1, a2, a3) can be calculated. Their values are as follows: </br></br> |
| - | <p align=center><img src="https://static.igem.org/mediawiki/2012/1/1f/ConsumptionRateConstants_university_of_british_columbia.png"></p> | + | <p align=center><img src="https://static.igem.org/mediawiki/2012/1/1f/ConsumptionRateConstants_university_of_british_columbia.png"width=600></p> |
| + | </br><b>Figure 4. Amino acid consumption measured from auxotroph cultures in minimial media spiked with an initial concentration of 0.15 mM of the required amino acid. Linear regression was used to find the following consumption rates: Trp: 0.304 ± 0.009 mM/OD (a1), Met: 0.115 ± 0.047 mM/OD (a2), Tyr: 0.143 ± 0.020 mM/OD (a3). Error bars represent 95% confidence interval as determined by regression in the calibration curves. </br></br> | ||
| + | <p align=left> | ||
Now that we have our consupmtion constants, we were interested in finding out how the other two auxotrophs will contribute to changes in environmental amino acid concentrations. </br></br> | Now that we have our consupmtion constants, we were interested in finding out how the other two auxotrophs will contribute to changes in environmental amino acid concentrations. </br></br> | ||
With regards to the behaviour of auxotrophs in coculture under changing amino acid concentrations, the closest assumption we can make is that each auxotroph will behave similarly to the conditions of monoculture. Thus, we used the same calibration curve for relating the final OD to the limiting substrate amino acid concentration in the media. The difference here is that, instead of finding out the corresponding limiting amino acid concentration, e.g., <i>ΔmetA</i> media for [Met] measurements, we are interested in measuring the other two non-limiting amino acid concentrations of each knocked out <i>E. coli</i> auxotrophs, e.g. <i>ΔmetA</i> media for [Trp] and [Tyr] concentrations, at various OD readings [2]. We were then able to derive six functions that relate the change of all amino acid concentrations with respect to time (e.g. d[Trp]/dt), based on two auxotrophs' optical densities (e.g. ODmetA<sup>-</sup> and ODtyrA<sup>-</sup>), as shown below: </br></br> | With regards to the behaviour of auxotrophs in coculture under changing amino acid concentrations, the closest assumption we can make is that each auxotroph will behave similarly to the conditions of monoculture. Thus, we used the same calibration curve for relating the final OD to the limiting substrate amino acid concentration in the media. The difference here is that, instead of finding out the corresponding limiting amino acid concentration, e.g., <i>ΔmetA</i> media for [Met] measurements, we are interested in measuring the other two non-limiting amino acid concentrations of each knocked out <i>E. coli</i> auxotrophs, e.g. <i>ΔmetA</i> media for [Trp] and [Tyr] concentrations, at various OD readings [2]. We were then able to derive six functions that relate the change of all amino acid concentrations with respect to time (e.g. d[Trp]/dt), based on two auxotrophs' optical densities (e.g. ODmetA<sup>-</sup> and ODtyrA<sup>-</sup>), as shown below: </br></br> | ||
| + | |||
| + | <p align=center><img src="https://static.igem.org/mediawiki/2012/f/f0/ODcalibrationTrp_university_of_british_columbia.png"></br><b> | ||
<p align=center><img src="https://static.igem.org/mediawiki/2012/b/b7/Equation10-15_data_analyzed_data_university_of_british_columbia.png"> | <p align=center><img src="https://static.igem.org/mediawiki/2012/b/b7/Equation10-15_data_analyzed_data_university_of_british_columbia.png"> | ||
| Line 84: | Line 77: | ||
Note: These equations can now be directly substituted into equations (1), (2), and (3), in order to simplify the Matlab Code section.</br></br> | Note: These equations can now be directly substituted into equations (1), (2), and (3), in order to simplify the Matlab Code section.</br></br> | ||
| - | + | <p align=left> | |
One shortcoming in the use of these equations is their dependency on the accuracy of the calibration curves (Figure 3, 4, and 5), as well as the OD readings, especially when OD readings are low and noisy (< 0.1, nearing the limits of the plate reader's accuracy). </br></br> | One shortcoming in the use of these equations is their dependency on the accuracy of the calibration curves (Figure 3, 4, and 5), as well as the OD readings, especially when OD readings are low and noisy (< 0.1, nearing the limits of the plate reader's accuracy). </br></br> | ||
| - | + | For example, one of the experimental raw data obtained are shown below: </br></br> | |
<p align=center><img src="https://static.igem.org/mediawiki/2012/2/2c/TyrA_in_metAknockoutssupernatants_university_of_british_columbia.png" width=800> | <p align=center><img src="https://static.igem.org/mediawiki/2012/2/2c/TyrA_in_metAknockoutssupernatants_university_of_british_columbia.png" width=800> | ||
| - | </br><b>Figure | + | </br><b>Figure 5: <i>ΔtyrA</i> auxotroph growth curve in various <i>ΔmetA</i> ODs cell free supernatants in 96-well plate reader environment. </b></br></br>. |
| + | <p align=left> | ||
Note: As discussed before, this may lead to inaccuracies in calculation from low OD readings. All data sets are labelled correspondingly, and triplicate measurements are noted by wells 1, 2, and 3. As we can see, there are some distinct differences in the OD readings among the three different supernatant data sets; however, when corrected by focusing on the difference between the initial reading and the final reading with a base adjustment of 0.025 (the lower value used for all the calibration curves), they average roughly the same, with an average OD of 0.03847, 0.04043, and 0.04218, respectively, for ODmet 0.219, 0.546, and 0.764. </br></br> | Note: As discussed before, this may lead to inaccuracies in calculation from low OD readings. All data sets are labelled correspondingly, and triplicate measurements are noted by wells 1, 2, and 3. As we can see, there are some distinct differences in the OD readings among the three different supernatant data sets; however, when corrected by focusing on the difference between the initial reading and the final reading with a base adjustment of 0.025 (the lower value used for all the calibration curves), they average roughly the same, with an average OD of 0.03847, 0.04043, and 0.04218, respectively, for ODmet 0.219, 0.546, and 0.764. </br></br> | ||
| Line 96: | Line 90: | ||
To illustrate this point more clearly, here is a comparison with another set of growth curve data run in the same plate.</br></br> | To illustrate this point more clearly, here is a comparison with another set of growth curve data run in the same plate.</br></br> | ||
<p align=center><img src="https://static.igem.org/mediawiki/2012/0/05/TyrA_and_trpA_in_metAknockoutssupernatants_university_of_british_columbia.png" width=800></br><b> | <p align=center><img src="https://static.igem.org/mediawiki/2012/0/05/TyrA_and_trpA_in_metAknockoutssupernatants_university_of_british_columbia.png" width=800></br><b> | ||
| - | Figure | + | Figure 6: Comparison between <i>ΔtrpA</i> and <i>ΔtyrA</i> auxotroph growth curve in the same supernatants of <i>ΔmetA</i> at its various optical density media.</p></b> |
| - | + | <p align=left> | |
Note that all three of the trpA<sup>-</sup> sets of growth curve grew to a much higher OD (reaching about 0.17) at the end of 15 hours and did not reach stationary phase, whereas all tyrA<sup>-</sup> sets grew significantly less, no significant change in OD at the end of the experiment.</br></br> | Note that all three of the trpA<sup>-</sup> sets of growth curve grew to a much higher OD (reaching about 0.17) at the end of 15 hours and did not reach stationary phase, whereas all tyrA<sup>-</sup> sets grew significantly less, no significant change in OD at the end of the experiment.</br></br> | ||
| Line 232: | Line 226: | ||
<p align=center><img src="https://static.igem.org/mediawiki/2012/c/c8/12hours_nomordificationprediction_university_of_british_columbia.png" width=700></br> | <p align=center><img src="https://static.igem.org/mediawiki/2012/c/c8/12hours_nomordificationprediction_university_of_british_columbia.png" width=700></br> | ||
| - | <b>Figure | + | <b>Figure 7. A comparison of model predictions versus empirical measurements (n=3) of each auxotroph’s growth in a co-culture.</br></br> |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | For our <b>future work</b>, we will refine our model with more experimental data. This can be accomplished by running the <a href="https://2012.igem.org/Team:British_Columbia/Protocols/AArate">amino acid</a> experiment data points of interest. Alternatively, we can use more direct methods of amino acid measurement, like HPLC analysis.</br></br> | + | <p align=left>As shown in the figure above, this model can give reasonable predictions though still can be further modified for better accuracy. For our <b>future work</b>, we will refine our model with more experimental data. This can be accomplished by running the <a href="https://2012.igem.org/Team:British_Columbia/Protocols/AArate">amino acid</a> experiment data points of interest. Alternatively, we can use more direct methods of amino acid measurement, like HPLC analysis.</br></br> |
This code can also be adapted to predict the population dynamics of our cocultures when tuning effects are introduced. This is accomplished by by generating six new functions to describe the amino acid dynamics of each monoculture. We can then use this model to decide the amount of tuning (by adding in different amount of inducer) needed to obtain a desired population of each auxotroph.</br></br> | This code can also be adapted to predict the population dynamics of our cocultures when tuning effects are introduced. This is accomplished by by generating six new functions to describe the amino acid dynamics of each monoculture. We can then use this model to decide the amount of tuning (by adding in different amount of inducer) needed to obtain a desired population of each auxotroph.</br></br> | ||
Latest revision as of 04:06, 26 October 2012

Modeling Microbial Consortia: The Auxotroph Approach
These data are fit to the Monod equation to determine the constants listed below.
Note: we used lower amino acid concentrations as well, however we found out that when concentration is below 1E-7 M, we observed no significant overnight growth of the cells. For a detailed protocol, please visit here.
Based on the fits presented in Figures 1 and 2, the key constants, such as maximum growth rates, and half-velocity constants, for each auxotroph were obtained [1]:
As discussed in the beginning of this section, we assumed that the amino acid consumption is solely dependent on the cell growth rate. In order to find out the consumption rate constants, (a1, a2, and a3), we need to find out a way to measure the amino acid concentrations in the media with respect to the change of its OD for each auxotroph. To do this, we need to find a way to measure media amino acid concentrations. With no good access to HPLC, we noticed in previous experiments that each of the knockout monocultures will reach to a certain final OD at a different known concentration of amino acid. In addition, recently published paper: ''A Programmable Escherichia coli Consortium via Tunable Symbiosis'', Kerner A et. al (2012) confirmed that there is indeed a relationship between the final auxotroph ODs with the environmental amino acid concentrations [2]. We thus generated a calibration curve to relate the limiting amino acid concentrations to their final OD readings. We based this on our own wet lab data shown in the figures below:
Figure 3: Calibration curves used to convert final OD600 to initial amino acid concentration. Standard solutions of known amino acid concentrations were innoculated with the respective auxotroph and grown for 20 hours at 37oC shaken at 200RPM, after which final OD600 were measured. Data were fit to quartic polynomials to inter- polate OD600 readings. Error bars represent standard deviation (n=3)..
We then sought out to find out how the amino acid in the environment is consumed with respect to cell growth.
The three auxotrophs were grown in 50 ml flasks with known initial Tryptophan, Methionine, and Tyrosine concentrations (5E-5 M), respectively, and 5ml of cell free supernatants at various ODs was isolated. These supernatants served as new 96-well plate media for each knock out cultures with newly added M9-Glucose shots (2% glucose in the well) to ensure the amino acids of interest remained as the limiting substrate for all knock outs. A detailed protocol can be found here.
The cells were grown in a 96-well plate for 15 hours at 37 oC, and growth curve data was obtained using a plate reader (most of the monocultures reached stationary phase at this point).
Based on the calibration curves (Figure 3), the amino acid concentrations in the wells were calculated from the final OD readings, and when corrected with a dilution factor of 5/4 (as we added in the 5X, it was 1/5 of the total well media volume, as described here), we obtained the resultant amino acid concentrations in the supernatant at various auxotroph monoculture densities. Again, with the assumption that the only change in the amino acid concentration in the media is due to cell growth, and as they are specific knock outs, they were not able to release any amino acid into the environment, each auxotrophs' consumption constants with respect to cell growth rate (a1, a2, a3) can be calculated. Their values are as follows:
Now that we have our consupmtion constants, we were interested in finding out how the other two auxotrophs will contribute to changes in environmental amino acid concentrations.
With regards to the behaviour of auxotrophs in coculture under changing amino acid concentrations, the closest assumption we can make is that each auxotroph will behave similarly to the conditions of monoculture. Thus, we used the same calibration curve for relating the final OD to the limiting substrate amino acid concentration in the media. The difference here is that, instead of finding out the corresponding limiting amino acid concentration, e.g., ΔmetA media for [Met] measurements, we are interested in measuring the other two non-limiting amino acid concentrations of each knocked out E. coli auxotrophs, e.g. ΔmetA media for [Trp] and [Tyr] concentrations, at various OD readings [2]. We were then able to derive six functions that relate the change of all amino acid concentrations with respect to time (e.g. d[Trp]/dt), based on two auxotrophs' optical densities (e.g. ODmetA- and ODtyrA-), as shown below:
One shortcoming in the use of these equations is their dependency on the accuracy of the calibration curves (Figure 3, 4, and 5), as well as the OD readings, especially when OD readings are low and noisy (< 0.1, nearing the limits of the plate reader's accuracy).
For example, one of the experimental raw data obtained are shown below:
Note: As discussed before, this may lead to inaccuracies in calculation from low OD readings. All data sets are labelled correspondingly, and triplicate measurements are noted by wells 1, 2, and 3. As we can see, there are some distinct differences in the OD readings among the three different supernatant data sets; however, when corrected by focusing on the difference between the initial reading and the final reading with a base adjustment of 0.025 (the lower value used for all the calibration curves), they average roughly the same, with an average OD of 0.03847, 0.04043, and 0.04218, respectively, for ODmet 0.219, 0.546, and 0.764.
To illustrate this point more clearly, here is a comparison with another set of growth curve data run in the same plate.
Note that all three of the trpA- sets of growth curve grew to a much higher OD (reaching about 0.17) at the end of 15 hours and did not reach stationary phase, whereas all tyrA- sets grew significantly less, no significant change in OD at the end of the experiment.
Matlab Code
Now with all the key functions and constants approximated, we are able to encode our model into Matlab, where the six key ODE functions can be solved and output the our prediction of population dynamics.
The code is shown below, which contains a detailed step by step instruction for future iGEM teams or related scientific researches use, as well as the clarifications of our own code. It is also written is a way that it can be directly copied and pasted into Matlab blank m.files with all the comments in Matlab commenting format already, so that the coder can read the coded instructions and explanations in the coding command window without having to switching between, for example, the matlab and browser windows.
Of course, copying and pasting the code here is welcomed!
Code begins here:
% input the following commands, which are all in bold, into MATLAB command window, and not the m.file, before you call the ODE function.
%firstly, define a time array, set the upper limit to be the time (hr) you want this model to predict. For this model to be accurate, it is advised not to use extreme values.
%in our cases, the upper limit is set to 12 as some of the modelling equations are based on 11 hour culture supernatant data. Thus accuracy is expected within 12 hours.
% xspan = [0 12];
%in order to set up the initial conditions, where they corespond to the initial values of y(1), y(2), y(3)..., to y(6) in our case, and y(n) if you have n number of ODE equations
% ynot = [4E-5 4E-5 4E-5 0.05 0.05 0.05]';
% the first three values are the initial values for y(1), y(2), and y(3) respectively, which in out cases are the initial amino acid concentrations, and as we are adding the cells directly into the M9 media without supplementing any amino acids, it is believed to be at a low concentrations, though there will be some left over amino acids within the living initial cells, and thus is set to 4E-5 mole/L.
% the remaining three values are the initial conditions for y(4), y(5), and y(6), which are the initial cell ODs in out case, and they are all set to 0.05, which is the initial OD reading we obtained after we innoculated the culture, with the assumption that all the cells are still alive.
%Then RUN THE FUNCTION
% [X,Y] = ode45(@UBCiGEM2012_ConsortiaModel,xspan,ynot);
% this takes the function name in your m.file, the range of your interest, and the initial conditions which is just defined, respectively
%Note: have to make sure that the name is exactly the same as the function name in your m.file for it to be able to run correctly.
%the following code is for the actual mfile.
% 1) Declare the name of the function:
function dy = UBCiGEM2012_ConsortiaModel(t,y);
% 2) define all your ODE variables clearly for your own understanding as they can get quite confusing if you don't.
% in our case:
% y(1) = [Trp];
% dy(1) = d[Trp]/dt;
% y(2) = [Met];
% dy(2) = d[Met]/dt;
% y(3) = [Tyr];
% dy(3) = d[Tyr]/dt;
% y(4) = [OD_trpA-];
% dy(4) = d[OD_trpA-]/dt;
% y(5) = [OD_tyrA-];
% dy(5) = d[OD_tyrA-]/dt;
% y(6) = [OD_metA-];
% dy(6) = d[OD_metA-]/dt;
% 3) define your constants first that will not depend on variables
% in our case, it was all measured through our own wet lab data and is explained in the previous section
%define maximum growth rate constants
u_maxtrpA = 0.23; % units = hr-1
u_maxmetA = 0.34; % units = hr-1
u_maxtyrA = 0.28; % units = hr-1
%define half-velocity constants
Ks_trpA = 3.30E-7; % units = M
Ks_metA = 5.10E-6; % units = M
Ks_tyrA = 8.50E-7; % units = M
%define the consumption rate constants
a1 = 3.05E-4; %units= M/OD trp
a2 = 1.15E-4; %units= M/OD met
a3 = 1.4E-4; %units= M/OD tyr
% 4) write out ODE CODE, suggest to write out all equations on paper first, as they are a lot more clear than coding them directly, and will reduce the chance for you to make error why coding significantly.
dy = zeros(6,1);
dy(1) = 2.5E-7 + 7.2E-8 - a1*(u_maxtrpA*(y(1))/(Ks_trpA+y(1)))*y(4);
dy(2) = 1.7E-7 + 5.1E-8 - a2*(u_maxmetA*(y(2))/(Ks_metA+y(2)))*y(5);
dy(3) = -3.6E-8 + 1.7E-6 - a3*(u_maxtyrA*(y(3))/(Ks_tyrA+y(3)))*y(6);
dy(4) = (u_maxtrpA*y(1))/(Ks_trpA+y(1))*(y(4));
dy(5) = (u_maxmetA*y(2))/(Ks_metA+y(2))*(y(5));
dy(6) = (u_maxtyrA*y(3))/(Ks_tyrA+y(3))*(y(6));
% note: For easy coding and checking purposes, all this equations are exactly the same as the one shown in equation (1), (2), and (3) for dy(1), dy(2), and dy(3) respectively, with also the same arrangements of each equation term within them.
% and Please also note: these equation has been derived with direct simple substitution of equations, where equation (4), (5), (6), (7), (8), (9), (10), (11), (12), (13), (14) and (15) have been substituted into the equation (1), (2) and (3).
%if you want, you can choose to plot diagrams and display the predicting dynamics within the m.file or manually plot the arrays of your interests in command window
%For example, for this project, we are interesting in predicting the ODs of each auxotrophs, thus, we manually plotted values in y(4), y(5), and y(6) (which again, are the OD values of each auxotrophs) agaisnt time, by typing the following in the command window after run the ODE45 functions:
%>>plot(X,Y(:,4:6))
End of the m.file
Simulation Results, Analysis and Future Work
When the Matlab Code was run, it generated a graphic result predicting the population dynamics without inducing, shown below:
As shown in the figure above, this model can give reasonable predictions though still can be further modified for better accuracy. For our future work, we will refine our model with more experimental data. This can be accomplished by running the amino acid experiment data points of interest. Alternatively, we can use more direct methods of amino acid measurement, like HPLC analysis.
This code can also be adapted to predict the population dynamics of our cocultures when tuning effects are introduced. This is accomplished by by generating six new functions to describe the amino acid dynamics of each monoculture. We can then use this model to decide the amount of tuning (by adding in different amount of inducer) needed to obtain a desired population of each auxotroph.
References
[1] Shuler, ML; Kargi, F; Bioprocess Engineering Basic Concepts, 2002 2nd edition. Prentice Hall PTR.
[2] Kerner A; Park J; Williams A; Lin XN; A Programmable Escherichia coli Consortium via Tunable Symbiosis, 2012. PLoS ONE 7(3): e34032. doi:10.1371/journal.pone.0034032
There is a large potential combinatorial space for optimizing and tuning a three-member microbial consortium with auxotrophic interdependence. In order to help address this concern, we set to develop a computational framework to help guide experimental decisions. Our model attempts to develop a predictive understanding of auxotrophic interdependence in a three-member microbial consortium. As it was important for us to be able to predict growth kinetics based on measurable, empirical properties, as well as to develop a mathematically and experimentally tractable system, we decided to base our model on Monod kinetics.
The Monod equation, models the growth of microorganisms in aqueous environments. The underlying assumption is that the growth rates are dependent on the concentration of a limiting nutrient. This equation relates the specific growth rate (µg, the specific growth rate of microorganisms) to the maximum growth rate (µm), the saturation constant (or half-velocity constant, Ks) as a function of the concentration of the limiting substrate (S) [1].
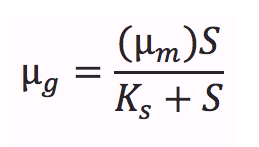
It takes into account a number of factors:
The following sections detail the mathematical foundation of the model, the experimental measurement of the necessary constants, the MATLAB implementation and some preliminary results.
Equations and Constants
We proposed the following fundamental equations for our consortium model. In this model, each auxotrophic strain is denoted with a superscript minus (i.e. ΔtrpA, ΔmetA, and ΔtyrA are denoted trpA-, metA-, and tryA-, respectively); [Trp], [Met], and [Tyr] represent the concentrations of tryptophan, methionine, and tyrosine; ODtrpA-, ODmetA-, and ODtyrA- represent the optical densities of the ΔtrpA, ΔmetA, and ΔtyrA strains; a1, a2, and a3 are the consumption constants with respect to changes in ODtrpA-, ODmetA-, and ODtyrA-, respectively; µtrpA-, µmetA-, µtyrA- are the specific growth rates of the ΔtrpA, ΔmetA, and ΔtyrA strains; µMaxtrpA-, µMaxmetA-, µMaxtyrA- are the maximum growth rates of the ΔtrpA, ΔmetA, and ΔtyrA strains; and KstrpA-, KsmetA-, KStyrA- are the half-velocity constants of the ΔtrpA, ΔmetA, and ΔtyrA strains. The terms and constants of this equation will be discussed in further detail later.

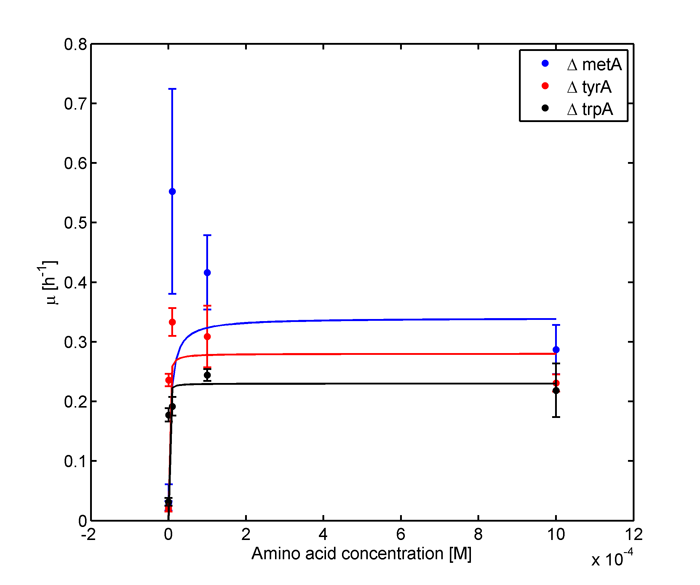 Figure 1: Monoculture Growth Rates at various Limiting Amino Acid Concentrations.
Figure 1: Monoculture Growth Rates at various Limiting Amino Acid Concentrations.
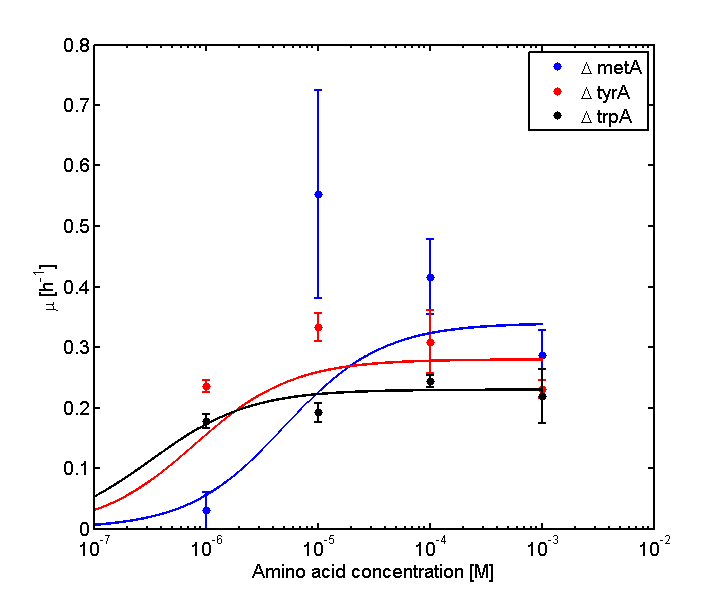 Figure 2: Log Scale of Monoculture Growth Rates at various Limiting Amino Acid Concentrations.
Figure 2: Log Scale of Monoculture Growth Rates at various Limiting Amino Acid Concentrations.
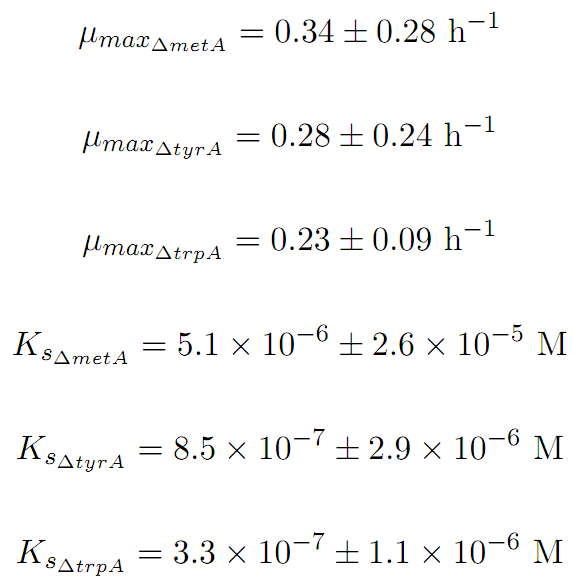
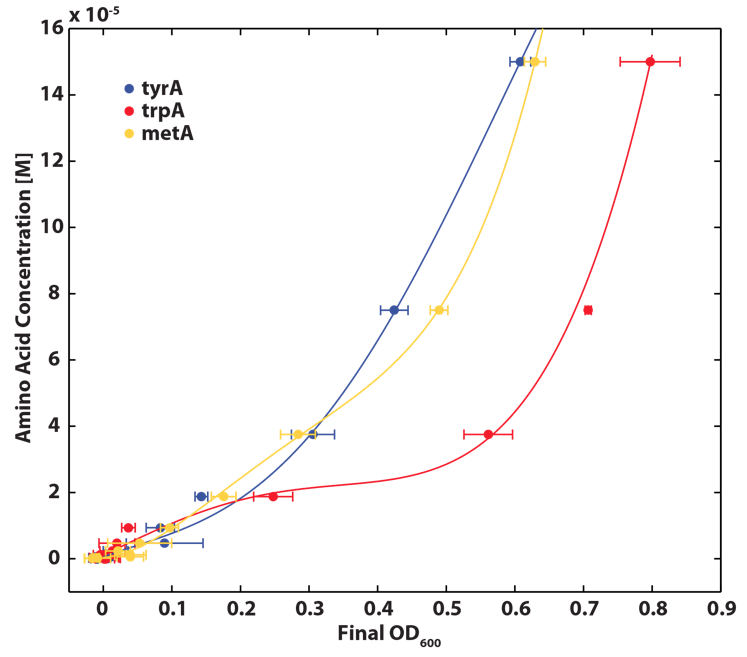
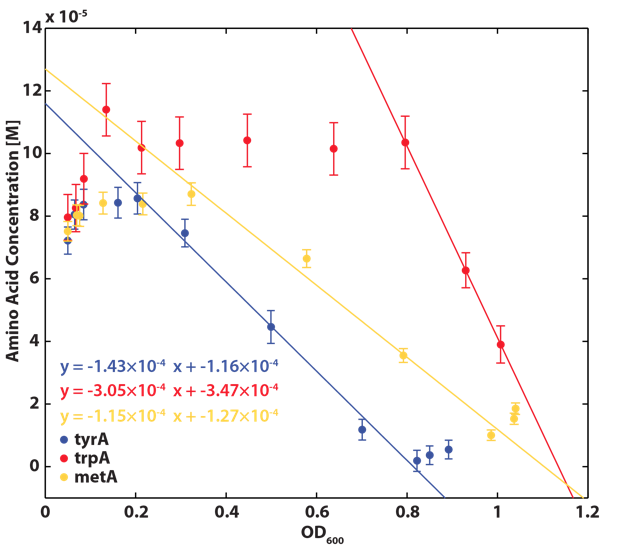
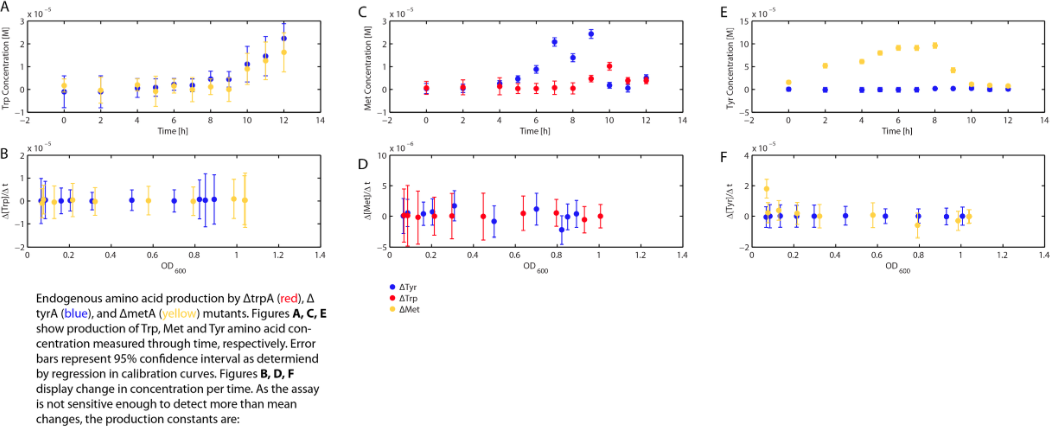

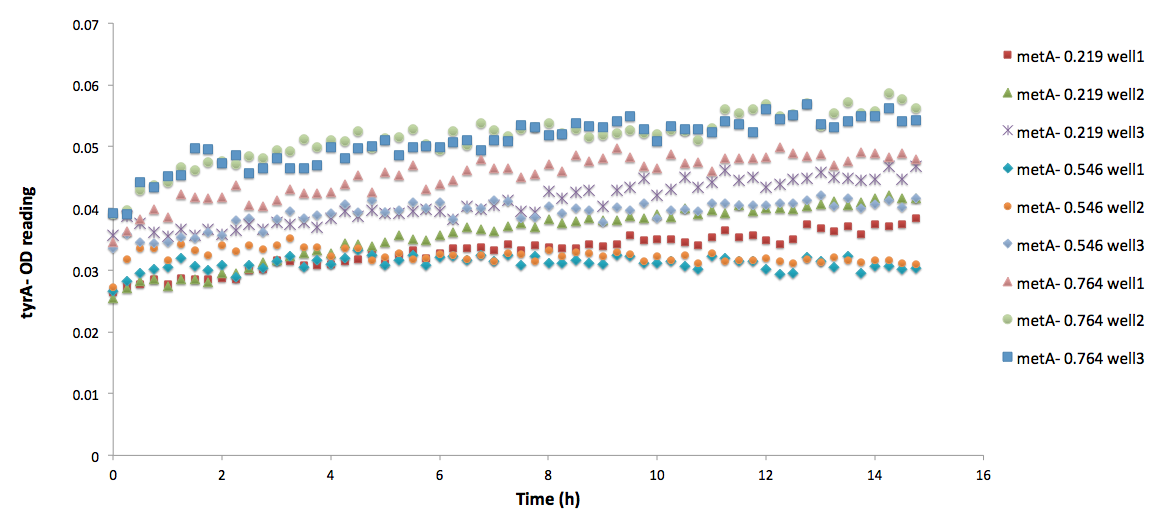 Figure 5: ΔtyrA auxotroph growth curve in various ΔmetA ODs cell free supernatants in 96-well plate reader environment. .
Figure 5: ΔtyrA auxotroph growth curve in various ΔmetA ODs cell free supernatants in 96-well plate reader environment. .
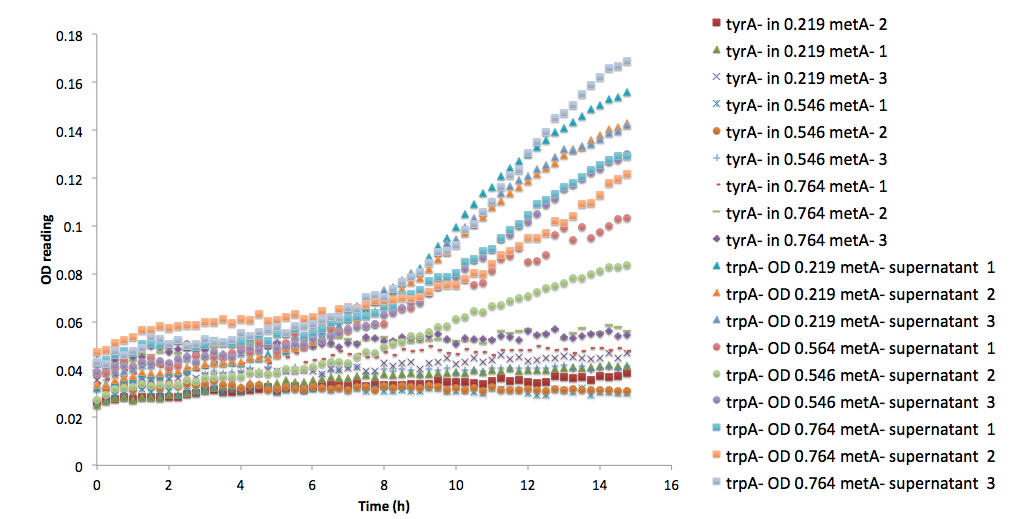 Figure 6: Comparison between ΔtrpA and ΔtyrA auxotroph growth curve in the same supernatants of ΔmetA at its various optical density media.
Figure 6: Comparison between ΔtrpA and ΔtyrA auxotroph growth curve in the same supernatants of ΔmetA at its various optical density media.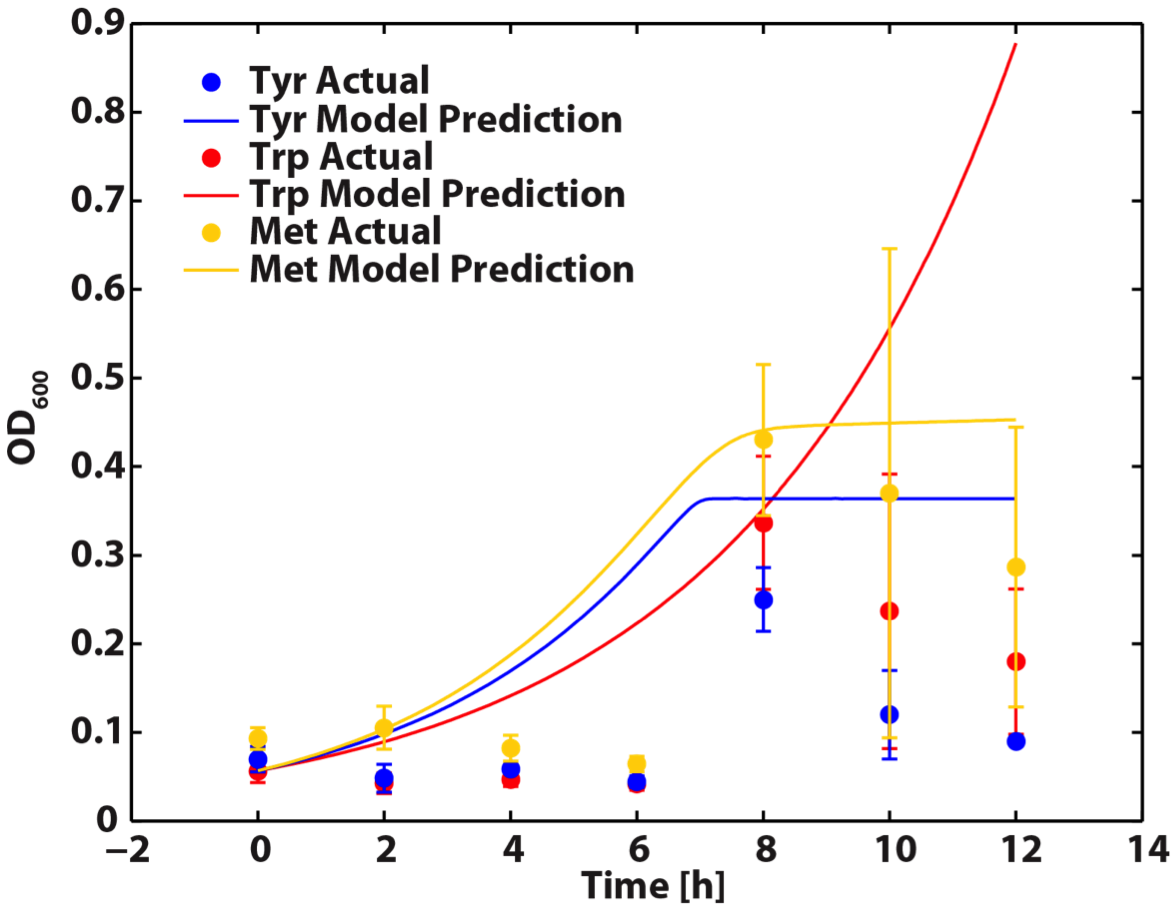 Figure 7. A comparison of model predictions versus empirical measurements (n=3) of each auxotroph’s growth in a co-culture.
Figure 7. A comparison of model predictions versus empirical measurements (n=3) of each auxotroph’s growth in a co-culture.