 "
"
Team:TU-Eindhoven/LEC/Modelling
From 2012.igem.org
PetraAlkema (Talk | contribs) |
|||
| Line 22: | Line 22: | ||
</p> | </p> | ||
<p> | <p> | ||
| - | The homeostatis system in yeast cells has <span class = "lightblue">two basic characteristics</span>. The cytosolic Ca<sup>2+</sup> concentration is tightly controlled by zero steady-state error to extracellular stimuli and the system is relatively insensitive to specific kinetic parameters, due to robustness of | + | The homeostatis system in yeast cells has <span class = "lightblue">two basic characteristics</span>. The cytosolic Ca<sup>2+</sup> concentration is tightly controlled by zero steady-state error to extracellular stimuli and the system is relatively insensitive to specific kinetic parameters, due to robustness of the adaptive response<html><a href="#ref_kitano" name="text_kitano"><sup>[2]</sup></a></html>. |
</p> | </p> | ||
| Line 48: | Line 48: | ||
</h3> | </h3> | ||
<p> | <p> | ||
| - | In literature, little can be found about modeling calcium channels in Saccaromyces Cerevisiae, most commonly known as budding yeast. Therefore we still used the model of <span class = "lightblue">sympathetic ganglion 'B' type cells of a bullfrog</span> to describe this process, since the type of voltage-dependent calcium channels is the same in both the bullfrog cells and the yeast cells. | + | In literature, little can be found about modeling calcium channels in <i>Saccaromyces Cerevisiae</i>, most commonly known as budding yeast. Therefore we still used the model of <span class = "lightblue">sympathetic ganglion 'B' type cells of a bullfrog</span> to describe this process, since the type of voltage-dependent calcium channels is the same in both the bullfrog cells and the yeast cells. |
</p> | </p> | ||
| Line 146: | Line 146: | ||
</p> | </p> | ||
<p> | <p> | ||
| - | The results, as presented in the previous section, show a large drop of the cytosolic Ca<sup>2+</sup>-level after removal of the potential difference. Although we could expect a fast recovery to the steady state, this drop is remarkably fast. This is one of the <span class = "lightblue">unexpected results</span> of this model and therefore one of the more interesting | + | The results, as presented in the previous section, show a large drop of the cytosolic Ca<sup>2+</sup>-level after removal of the potential difference. Although we could expect a fast recovery to the steady state, this drop is remarkably fast. This is one of the <span class = "lightblue">unexpected results</span> of this model and therefore one of the more interesting features to validate with experiments. Especially since this drop will influence the concentration of calcium bounded GECO-protein. |
</p> | </p> | ||
<p> | <p> | ||
| Line 158: | Line 158: | ||
<p> | <p> | ||
| - | To examine the calcium in the cell, a dynamic calcium model was designed. As a start a <span class = "lightblue">basic model of calcium homeostasis in yeast cells</span> was presented and tested. This model was extended | + | To examine the calcium in the cell, a dynamic calcium model was designed. As a start a <span class = "lightblue">basic model of calcium homeostasis in yeast cells</span> was presented and tested. This model was extended to include overexpression of voltage-dependent calcium channels and addition of GECO-kinetics. With this model, we can <span class = "lightblue">test and verify theoretical hypotheses</span> by comparing simulation results with corresponding experimental results and generate new hypotheses on the regulation of calcium homeostasis. On the other hand, due to the existence of unknown kinetic parameters and the lack of experimental data, this model is <span class = "lightblue">not an exact model yet</span>. However, it does can give some quantitative insight into the possible dynamics of the whole process and provide a <span class = "lightblue">general framework</span> for more elaborate investigations. |
</p> | </p> | ||
Revision as of 20:45, 25 September 2012

Global introduction
Biological cells use highly regulated homeostasis systems to keep a very low cytosolic Ca2+ level. In normal-growing yeast the cytosolic Ca2+ concentration is maintained in the range of 50-200 nM in the presence of environmental Ca2+ concentrations ranging from µM to 100 mM[1].
The homeostatis system in yeast cells has two basic characteristics. The cytosolic Ca2+ concentration is tightly controlled by zero steady-state error to extracellular stimuli and the system is relatively insensitive to specific kinetic parameters, due to robustness of the adaptive response[2].
To achieve an accurate model, the influences of voltage-dependent calcium channels are added to a basic model for yeast calcium homeostasis. In this model, first described by J. Cui et al, the main contributions of calcium transport are defined[3].
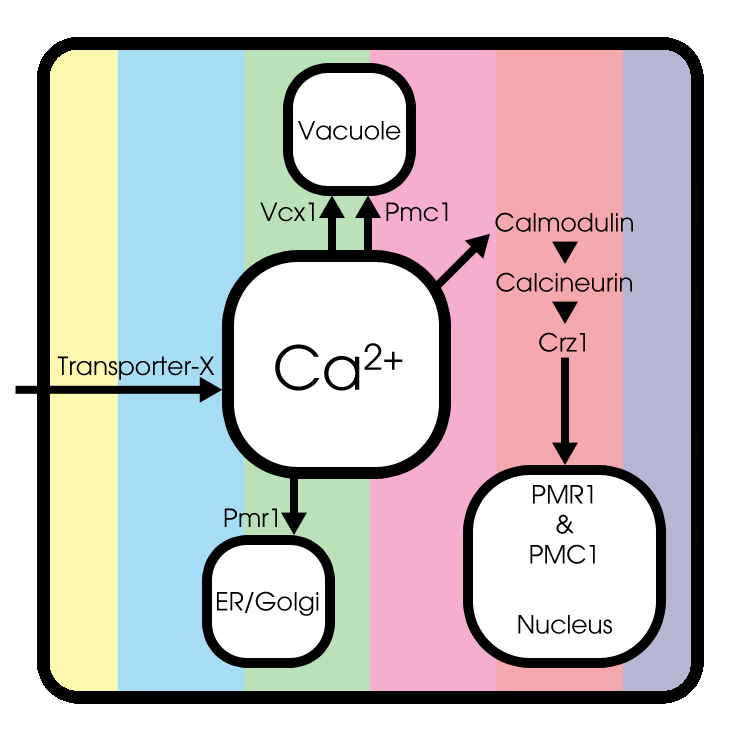
Basic model
Under normal conditions, extracellular Ca2+ enters the cytosol through an unknown Transporter X, whose encoded gene has not been identified yet. Cytosolic Ca2+ can be pumped into the endoplasmic reticulum (ER) and Golgi system through Pmr1 and can be sequestered into the vacuole through Pmc1 and Vcx1.
Both the expression and function of Pmc1, Pmr1 and Vcx1 are regulated by calcineurin, a highly conserved protein phosphatase that is activated by Ca2+-bound calmodulin[3]. Therefore, the transcription factor Crz1 can be dephosporylated by activated calcineurin. A conformational switch model is used to simulate Crz1 translocation, as described by Okamuraet al [4].
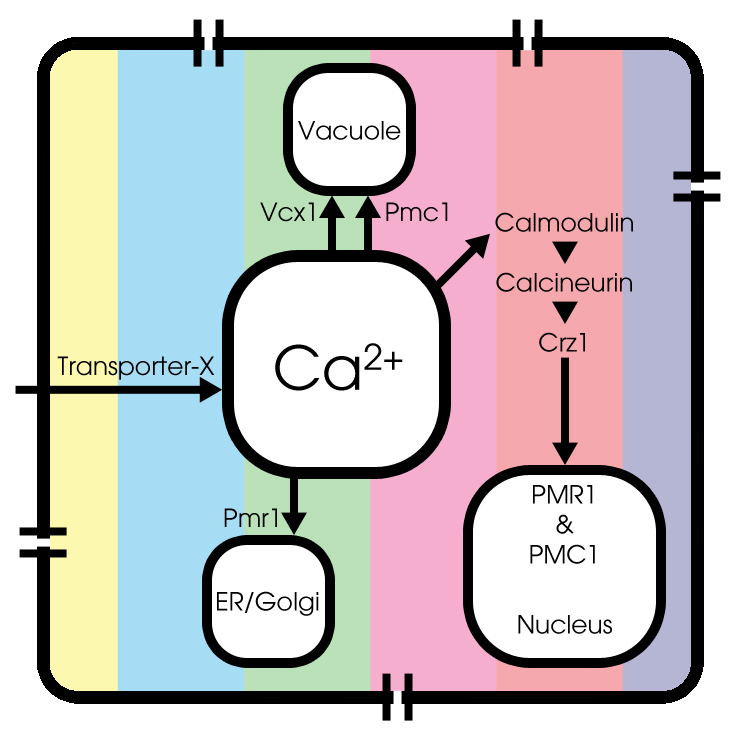
Voltage-dependent calcium channels
In literature, little can be found about modeling calcium channels in Saccaromyces Cerevisiae, most commonly known as budding yeast. Therefore we still used the model of sympathetic ganglion 'B' type cells of a bullfrog to describe this process, since the type of voltage-dependent calcium channels is the same in both the bullfrog cells and the yeast cells.
More information on the voltage-dependent calcium channels is presented in the description of the inital model.
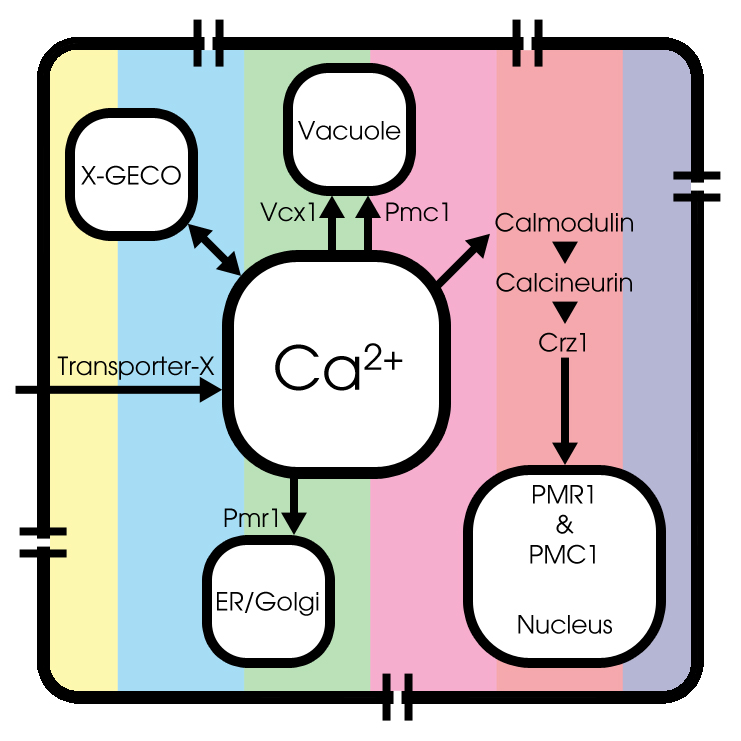
GECO-proteins
Last, but certainly not least, we added the kinetics of calcium binding by the GECO-proteins, since we use the fluorescent activity of CMV-X-GECO.1 proteins as light-emitting part of the cell, as a result of an increase of the cytosolic Ca2+-level.
The kinetic characterization of the GECO-proteins:

(Click to enlarge)
In short, we formulated a new basic calcium model for yeast cells. This model is extended with overexpression of voltage-dependent calcium channels and addition of GECO-kinetics. The model is made in MATLAB and the code can be found here.
In more detail
Since we do not want to bother the semi-interested readers with the complete derivation of the differential equations describing this model, this can be found in the bachelor thesis of one of our team members, Petra Alkema. In order to understand the results, discussion and conclusion in more detail, we really recommend to study this chapter. In an addition, a more detailed introduction to the conformational switch model, part of the total model, can be found here.
The main differential equations are:

Parameters
The model parameters for which all results are calculated, unless otherwise stated:

Results
As we could not yet accomplish a complete sensitivity analysis on the model, we only consider the basic characteristics of the different parts of the model. Furthermore, the influences of some basic experimental setup values could be stated, i.e. the concentration of extracellular calcium, Ca{ex}, and the duration of the pulse. You can click on the figures to enlarge the results. The first two graphs present the basic model in yeast cells, with the expected oscillatory system. The next graphs show the behavior of the complete model, including the voltage pulse. More results and a more detailed description can be found in the thesis.




Discussion
One of the most important points of discussion is the overall correctness of the calcium model. Due to the fact that the characteristics of both the voltage-dependent calcium channels and the GECO-proteins are added to the calcium homeostasis of the organism Saccaromyces Cerevisiae, a completely new system is created. As far as we know, in literature, no experimental data can be found about this specific system. At the moment of writing, the wetwork of the iGEM team of Eindhoven University of Technology 2012 is unfortunately not able to provide applicable information. A great step forward could be made when this model can be checked with experimental data. Nevertheless this model does give some quantitative insight in the behavior.
The simplification of the calcium homeostasis in a living yeast cell certainly leads to some imperfections upon the level of the actual physiology. For example, in the current model, we assume that the concentrations of Pmc1 and Pmr1 are directly proportional to the quantity of transcriptionally active Crz1. This is a big simplification since in real cells, this process involves the increased gene expression through Crz1 followed by translation and transport of the proteins to the respective intracellular destinations. Moreover, the current model does not include the influence from other relevant pathways whereas in real cells, any response to given extracellular stimulus is likely to be the result of complex cross-talk between multiple pathways.
Also some other assumptions need to be taken into account. First, we assumed that only fully dephosphorylated Crz1 molecules in the nucleus are transcriptionally active since this has been shown the case for NFAT1. Although the mechanism of Crz1 translocation in yeast cells is strikingly similar to NFAT, not all processes can be regarded the same, since there are no experimental data found to validate this assumption. Second, we assumed the behavior of the voltage-dependent calcium channels in yeast cells to be the same as in sympathetic ganglion 'B' type cells of a bullfrog.
The results, as presented in the previous section, show a large drop of the cytosolic Ca2+-level after removal of the potential difference. Although we could expect a fast recovery to the steady state, this drop is remarkably fast. This is one of the unexpected results of this model and therefore one of the more interesting features to validate with experiments. Especially since this drop will influence the concentration of calcium bounded GECO-protein.
In order to provide more quantitative insights in the experimental setup for the total project, we have to obtain information about the fluorescent properties of the GECO-proteins. At this moment we can only predict when the CaGECO concentration reaches a set percentage of the maximum amount of Ca2+-bounded GECO-protein. An important value is the amount of calcium bounded GECO-protein that needs to be there in order to detect fluorescent light.
Conclusion
To examine the calcium in the cell, a dynamic calcium model was designed. As a start a basic model of calcium homeostasis in yeast cells was presented and tested. This model was extended to include overexpression of voltage-dependent calcium channels and addition of GECO-kinetics. With this model, we can test and verify theoretical hypotheses by comparing simulation results with corresponding experimental results and generate new hypotheses on the regulation of calcium homeostasis. On the other hand, due to the existence of unknown kinetic parameters and the lack of experimental data, this model is not an exact model yet. However, it does can give some quantitative insight into the possible dynamics of the whole process and provide a general framework for more elaborate investigations.
We really hope that in the future of the iGEM competition one of the new iGEM teams will adapt our modeling project of calcium dynamics in yeast cells! :-)
References
- [1] A. Miseta, L. Fu, R. Kellermayer, J. Buckley, D.M. Bedwell, “The Golgi Apparatus plays a significant role in the maintainance of Ca2+ homeostasis in the vps33 vacuolar biogenesis mutant of Saccharomyces cerevisiae”, J. Biol. Chem. 274: 5939-5947, (1999)
- [2] H. Kitano, Systems biology : a brief overview, Nature 295: 1662-1664, (2002)
- [3] J. Cui, Mathematical modeling of metal ion homeostasis and signaling systems, (2009)
- [4] H. Okamura, J. Aramburu, C. Garca-Rodrguez, et al. Concerted dephosphorylation of the transcription factor NFAT1 induces a conformational switch that regulates transcriptional activity, Mol. Cell 6: 539-550, (2000)