 "
"
Team:Cornell/project/wetlab/chassis
From 2012.igem.org
(Difference between revisions)
R.Lizarralde (Talk | contribs) |
|||
| (4 intermediate revisions not shown) | |||
| Line 37: | Line 37: | ||
</li> | </li> | ||
<li> | <li> | ||
| - | <a href="https://2012.igem.org/Team:Cornell/project/wetlab/results/ | + | <a href="https://2012.igem.org/Team:Cornell/project/wetlab/results/transcription">Transcriptional Characterization</a> |
</li> | </li> | ||
<li> | <li> | ||
| - | <a href="https://2012.igem.org/Team:Cornell/project/wetlab/results/ | + | <a href="https://2012.igem.org/Team:Cornell/project/wetlab/results/currentresponse">Current Response</a> |
</li> | </li> | ||
| - | |||
<li> | <li> | ||
<a href="https://2012.igem.org/Team:Cornell/project/wetlab/results/protein">MtrB Protein Expression</a> | <a href="https://2012.igem.org/Team:Cornell/project/wetlab/results/protein">MtrB Protein Expression</a> | ||
| Line 76: | Line 75: | ||
</div> | </div> | ||
<div class="three columns"> | <div class="three columns"> | ||
| - | <a href="https://static.igem.org/mediawiki/2012/0/08/SEMMtrABCL.png" rel="lightbox"><img src="https://static.igem.org/mediawiki/2012/ | + | <a href="https://static.igem.org/mediawiki/2012/0/08/SEMMtrABCL.png" rel="lightbox"><img src="https://static.igem.org/mediawiki/2012/d/d3/SEMMtrABCLs.png"></a> |
</div> | </div> | ||
</div> | </div> | ||
<div class="row" id="video" style="margin:10px auto;"> | <div class="row" id="video" style="margin:10px auto;"> | ||
| - | <iframe width=" | + | <iframe width="765" height="465" src="http://www.youtube.com/embed/OSNLX-DI_kQ" frameborder="0" allowfullscreen></iframe> |
</div> | </div> | ||
<div class="row" id="item2"> | <div class="row" id="item2"> | ||
| Line 169: | Line 168: | ||
<script src="https://2012.igem.org/Team:Cornell/javascripts/foundation.min?action=raw&ctype=text/javascript"></script> | <script src="https://2012.igem.org/Team:Cornell/javascripts/foundation.min?action=raw&ctype=text/javascript"></script> | ||
<script src="https://2012.igem.org/Team:Cornell/javascripts/app?action=raw&ctype=text/javascript"></script> | <script src="https://2012.igem.org/Team:Cornell/javascripts/app?action=raw&ctype=text/javascript"></script> | ||
| + | <script src="https://2012.igem.org/Team:Cornell/javascripts/lightbox?action=raw&ctype=text/javascript"></script> | ||
<script type="text/javascript"> | <script type="text/javascript"> | ||
$(window).load(function() { | $(window).load(function() { | ||
Latest revision as of 00:25, 27 October 2012








Chassis
Introduction
Shewanella oneidensis MR-1 is a gram-negative facultative anaerobe capable of transferring electrons from the cytoplasmic space to electron acceptors outside the outer membrane (OM) [1,2]. In particular, the Mtr respiratory pathway of MR-1 is required for the extracellular reduction of metals and metal oxides 2. The Shewanella Mtr gene cluster encodes three outer membrane cytochromes (OMCs), MtrF, OmcA, and MtrC, two periplasmic decaheme cytochromes, MtrD and MtrA, and two alleged OM b-barrel proteins, MtrB and MtrE. The current model of Mtr respiration begins with electron transfer from carbon source catabolism to CymA, a cytoplasmic membrane (CM) cytochrome protein via the menaquinone pool. From there, electrons can be transferred to a number of different periplasmic electron carriers (PECs). MtrA, a decaheme c-type cytochrome and the predominant PEC for reduction of iron(III) and flavins, is physically associated with MtrB, a b-barrel protein spanning the outer membrane. Together, the MtrAB complex transfers electrons to OmcA and MtrC, two outer-membrane c-type cytochromes, which can function as the terminal reductases for a number of different metal substrates [1,4].

CymA
CymA, a c-type tetraheme cytochrome protein located in the CM, belongs to the NapC/NrfH family of quinol dehydrogenases principal in bacterial anaerobic respiration [4]. CymA is anchored in the cytoplasmic membrane by a single transmembrane domain with the rest of the protein extending into the periplasmic space; four heme groups are covalently bound to the periplasmic portion of the CymA protein. CymA has been shown to be essential to the reduction of Fe(III), DMSO, fumarate, nitrate, manganese and nitrite in Shewanella species through its shuttling of electrons from the cytoplasm to a number of different PECs [4,5]. Whereas NrfH proteins form strong bonds with their redox partners, CymA appears to bond only transiently to its partners in the periplasm. This is consistent with its ability to interact with many different PECs, offering Shewanella sp. respiration flexibility and versatility [4].MtrA
Reverse electron flow experiments have shown that direct electron transfer occurs between CymA and MtrA, a periplasmic decaheme c-type cytochrome, during in vitro Fe(III) oxide reduction [4]. Recently, the structure of MtrA has been elucidated through the use of small angle x-ray scattering and analytical ultracentrifugation; due to its elongated and 104 Å rod-like shape, MtrA has been postulated to function as an electron shuttle between CymA and OMCs MtrC or OmcA [2,3]. Cross-linking by formaldehyde treatment has shown that MtrA is physically associated with MtrB [7].
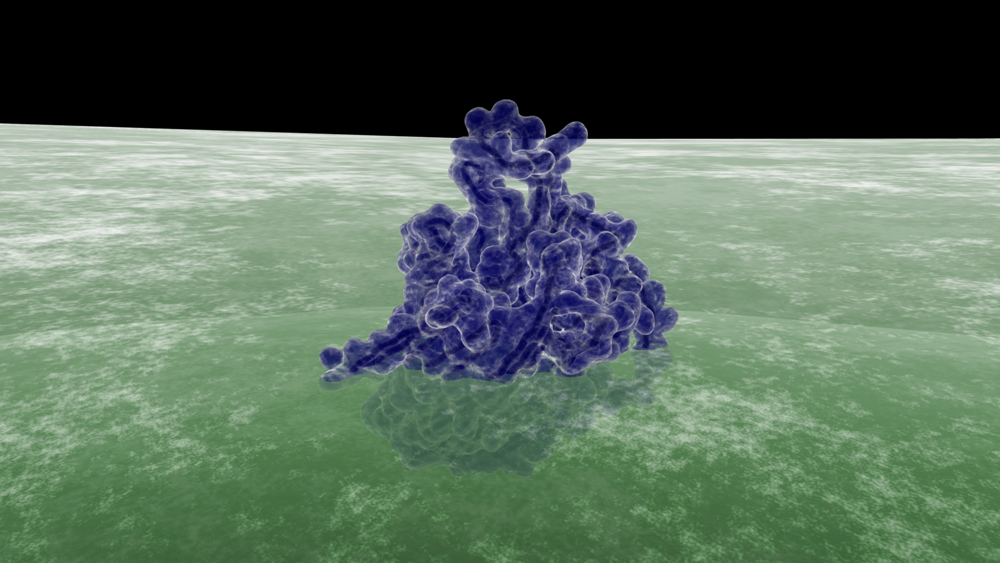
MtrB
While the structure of MtrB has not been determined, based on sequence analysis, MtrB has been posited to be an OM b-barrel, porin-like protein consisting of 28 transmembrane b-strands [1,4,7]. Experiments done with mtrB deletion mutants have shown that MtrB is required for MtrA localization to the outer membrane. Additionally, in a mtrA deletion strain, mtrB expression is detected in cell lysates but MtrB is not, indicating the necessity of MtrA in assembly of OM MtrB complexes [7]. This tight interaction between MtrA and MtrB has raised the possibility that MtrA may be embedded within MtrB, thereby allowing MtrA to come in contact with OM cytochrome proteins. Given the periplasmic length of approximately 237 Å and the localization of MtrA and CymA to the OM and CM membranes respectively, it is hypothesized that additional PECs, likely MtrA or NrfA paralogs, are necessary to complete the transfer of electrons between CymA and MtrA in MR-1 [7].MtrC and OmcA
MtrC and OmcA are two c-type decaheme cytochromes located on the extracellular surface of the OM [4,7]. In vivo cross-linking and in vitro experiments have shown that these two OMC readily complex and that this binding synergistically increases the reductase efficiency of the complex [7]. The current model of MtrC-OmcA metal reduction posits that MtrC and OmcA bind and reduce Fe(III) oxides directly [7]. Study of MtrF, a MtrC paralog, has provided structural evidence supporting the role of MtrC and OmcA as the terminal reductases in Fe(III) oxide reduction. MtrF folds into four domains with domains II and IV, the pentaheme domains, forming a core. Two split-b-barrels, domains I and III, surround the pentaheme cores [4]. Domains I and III are thought to be responsible for binding and reducing Fe(III), domain II for the direct transfer of electrons, and domain IV, for physically interacting with the MtrDE (a MtrAB homolog) complex [4].
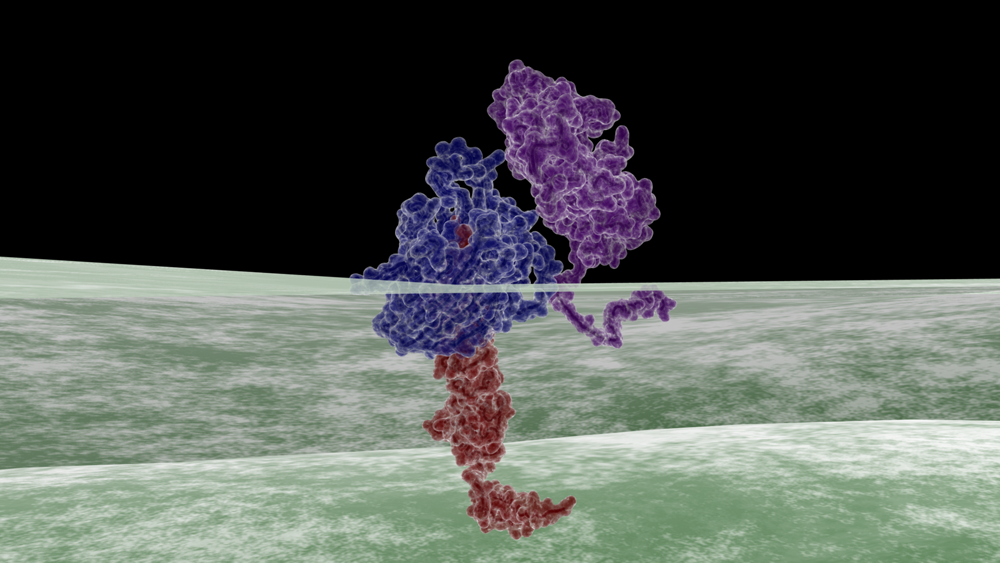
The Essential Role of MtrB in the Mtr Respiratory Pathway
In addition to serving as a conduit for electron transfer from MtrA to OM reducatses, MtrB acts as a scaffolding protein. MtrABC complexes are readily isolated after Triton X-100 treatment [7,8]. Isolated MtrABC, when solubilized into liposomes, readily catalyzes transmembrane electron transfer [7]. While MtrAB complexes readily form in the absence of MtrC, MtrBC complexes are not detected in a mtrA deletion background 7. Moreover, while isolated MtrA and MtrC can complex, it appears that MtrB is further needed in order for MtrAC interactions to be stable [7]. This indicates that in vivo, there is likely a requirement for association of MtrA and B, followed by association with MtrC for efficient electron transfer. MtrB is also integral to the correct localization of MtrC, MtrA, and OmcA to their respective positions along the Mtr respiratory pathway. In a mtrB deletion background, MtrA is unable to localize to the OM [7]; similarly, significant amounts of OmcA and MtrC have been shown to mislocalize to the CM in mtrB deletion strains [9]. Of the b-barrel mtrB paralogs that exist in the Shewanella genome, mtrB has been shown to be the most functional in Fe(III) citrate reduction. Coursolle and Gralnick, 2012, recently looked at different combinations of PECs, b-barrel, and OMC paralogs using a DMtr/DmtrB/mtrE strain. Of the functional iron-reducing modules examined, MtrA/MtrB/MtrC showed the highest Fe(III) citrate reduction levels followed by MtrA/MtrB/MtrF . Combinations with MtrE, a MtrB paralog, were also able to reduce Fe(III) citrate, albeit at a lower rate, while neither DmsF nor S04359 produced functional iron citrate reducing modules [1]. This combined with the integral role of MtrB in positioning of PEC and OMCs along the Mtr respiratory pathway makes it an ideal target for our reporter systems.References
1. Coursolle, D., & Gralnick, J. a. (2012). Reconstruction of Extracellular Respiratory Pathways for Iron(III) Reduction in Shewanella Oneidensis Strain MR-1. Frontiers in microbiology, 3(February), 56. doi:10.3389/fmicb.2012.000562. Coursolle, D., Baron, D. B., Bond, D. R., & Gralnick, J. A. (n.d.). The Mtr Respiratory Pathway Is Essential for Reducing Flavins and Electrodes in Shewanella oneidensis . Journal of Bacteriology , 192 (2 ), 467-474. doi:10.1128/JB.00925-09
3. Firer-Sherwood, M. A., Ando, N., Drennan, C. L., & Elliott, S. J. (2011). Solution-Based Structural Analysis of the Decaheme Cytochrome, MtrA, by Small-Angle X-ray Scattering and Analytical Ultracentrifugation. The Journal of Physical Chemistry B, 115(38), 11208-11214. American Chemical Society. doi:10.1021/jp203603r
4. Shi L, Rosso KM, Clarke TA, Richardson DJ, Zachara JM and Fredrickson JK (2012) Molecular underpinnings of Fe(III) oxide reduction by Shewanella oneidensis MR-1. Front. Microbio. 3:50. doi: 10.3389/fmicb.2012.00050
5. Myers, J. M., & Myers, C. R. (2000). Role of the Tetraheme Cytochrome CymA in Anaerobic Electron Transport in Cells of Shewanella putrefaciens MR-1 with Normal Levels of Menaquinone . Journal of Bacteriology , 182 (1 ), 67-75. doi:10.1128/JB.182.1.67-75.2000
6. Bretschger, O., Obraztsova, A., Sturm, C. A., Chang, I. S., Gorby, Y. A., Reed, S. B., Culley, D. E., et al. (n.d.). Current Production and Metal Oxide Reduction by Shewanella oneidensis MR-1 Wild Type and Mutants . Applied and Environmental Microbiology , 73 (21 ), 7003-7012. doi:10.1128/AEM.01087-07
7. Hartshorne, R. S., Reardon, C. L., Ross, D., Nuester, J., Clarke, T. A., Gates, A. J., Mills, P. C., et al. (2009). Characterization of an electron conduit between bacteria and the extracellular environment . Proceedings of the National Academy of Sciences . doi:10.1073/pnas.0900086106
8. Pitts, K. E., Dobbin, P. S., Reyes-Ramirez, F., Thomson, A. J., Richardson, D. J., & Seward, H. E. (2003). Characterization of the Shewanella oneidensis MR-1 Decaheme Cytochrome MtrA . Journal of Biological Chemistry , 278 (30 ), 27758-27765. doi:10.1074/jbc.M302582200
9. Shi, L., Chen, B., Wang, Z., Elias, D. A., Mayer, M. U., Gorby, Y. A., Ni, S., et al. (n.d.). Isolation of a High-Affinity Functional Protein Complex between OmcA and MtrC: Two Outer Membrane Decaheme c-Type Cytochromes of Shewanella oneidensis MR-1 . Journal of Bacteriology , 188 (13 ), 4705-4714. doi:10.1128/JB.01966-05