Membrane Accelerator - Fatty Acid and Alkane Biosynthesis
Fatty acids are promising biofuel precursors with a broad and bright future. An enzyme system of nine proteins is responsible for fatty acid biosynthesis in E.coli and is first introduced to iGEM by our team. We choose four crucial enzymes, TesA, FabG, FabZ, and FabI, to prove the feasibility of the Membrane Accelerator and to enhance rate of fatty acids synthesis. Moreover, we tried to further accelerate the alkane biosynthetic pathway in E.coli.
To identify and evaluate two outstanding features of the Membrane as a scaffold: the priority of products to exportation and the refinement of interaction.
To testify the feasibility of the Membrane Accelerator device in the context of fatty acids synthesis.
To upgrade fatty-acid-producing Membrane Accelerator device with alkane synthesis function.
To justify the universality of Membrane Accelerator
We constructed four enzyme-bearing fusion proteins that localize on membrane and assemble orderly.
We increased the yield of fatty acids by 50% only by localizing single TesA onto E.coli's inner membrane.
We successfully increased the yield of fatty acids by 24 fold by localizing TesA, FabG, FabZ, and FabI onto E.coli's inner membrane.
We upgraded fatty-acid-producing Membrane Accelerator by introducing additional two enzymes that convert fatty acids into alkane.
Background
We put Membrane Accelerator system into practice context to justify its versatility and efficiency.
Fatty acid derivatives are to emerge as prominent fuels in the marketplace. Their high energy density and low water solubility make them promising alternatives to fossil fuel as transportation fuels. Biosynthesis, among all the biofuels production methods, is the most sustainable, environmental friendly and less controversial one.
Initial fatty acid biosynthesis in E.coli is catalyzed by a series of nine enzymes and the final release of free fatty acids is catalyzed by a thioesterase via hydrolysis of acyl-ACP species. We are the first group in iGEM competition to recruit this biosynthetic pathway. Membrane Accelerator Device is expected to facilitate the biosynthesis of fatty acid in E.coli. Furthermore, we tried to upgrade fatty-acid-producing Membrane Accelerator by introducing additional two enzymes that convert fatty acids into alkane. Thus, alkane would be efficiently synthesized in E.coli directly.
Biosynthetic Pathway
Fatty acid biosynthetic pathway
Carbon chain elongation of fatty acid in E.coli is carried out by a nine-component enzyme system , FabA, FabB, FabD, FabF, FabG, FabH, FabI, FabZ, and ACP. They orderly cooperate to convert one equivalent of acetyl-CoA and 6-8 equivalents of malonyl-CoA into C14-C18 acyl-ACP species. The cytoplasmic mutant of the periplasmic thioesterase is capable of releasing free fatty acids preventing the fatty acyl yields from being directly harnessed for phospholipid biosynthesis.
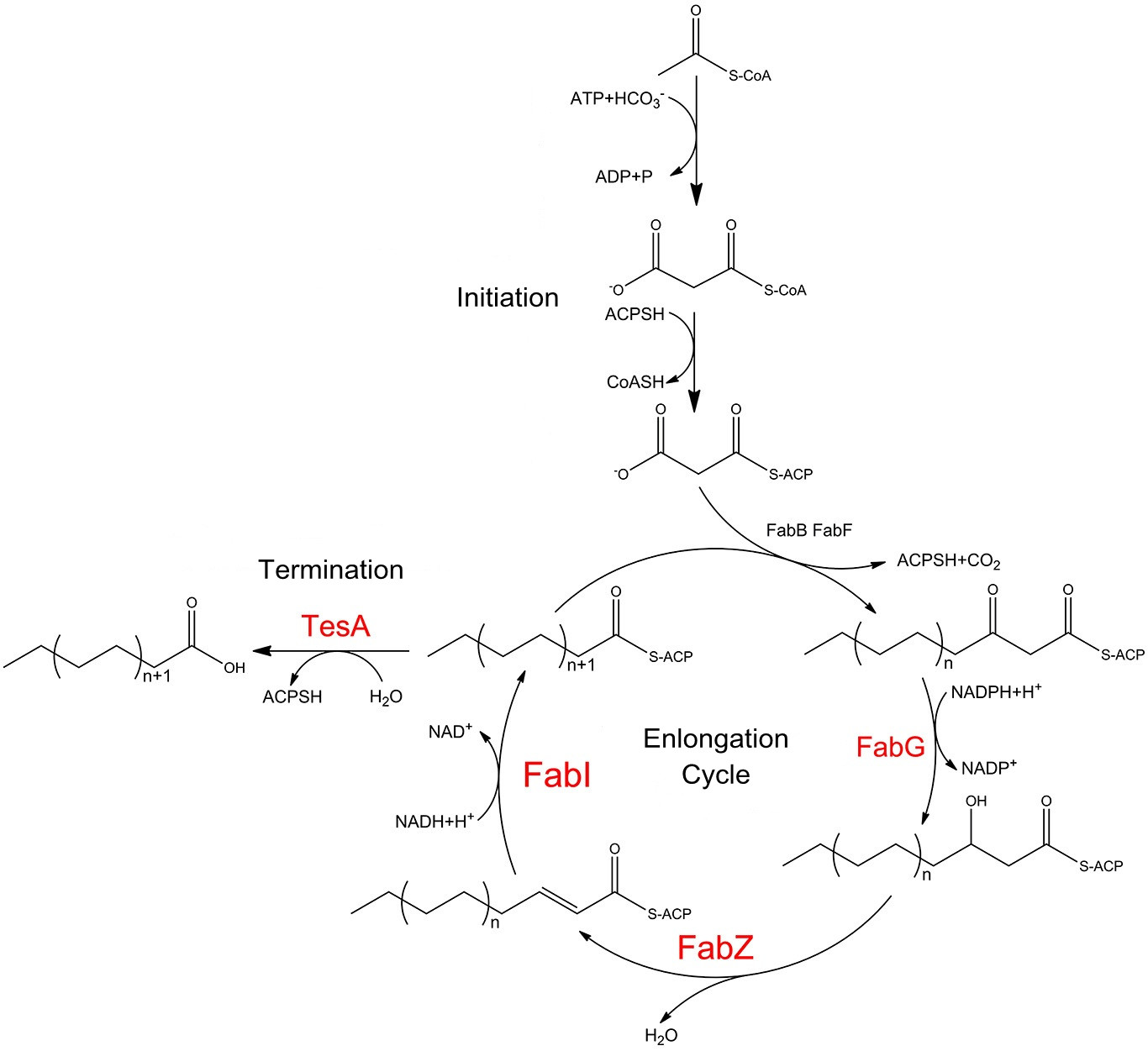 Fig.1 shows fatty acid biosynthesis pathways in E.coli, stressing the role of FabG, FabZ, FabI and TesA. Catalytic cycle of the E. coli fatty acid synthesis is initiated when holo-ACP, NADPH and NADH, acetyl-CoA and malonyl-CoA undergo condensation and subsequent reduction to form butyryl-ACP. These reactions are catalyzed by the malonyl-CoA:ACP transacylase FabD, the ketosynthase FabH, the NADPH-dependent ketoreductase FabG, either the dual-function dehydratase/isomerase FabA or the monofunctional dehydratase FabZ, and the NADH-dependent enoyl reductase FabI. Then the butyryl-ACP is extended via 5-7 rounds of analogous reactions to produce a C14 to C18-ACP either fully saturated or monounsaturated. These extension cycles are catalyzed by either the ketosynthase FabB or FabF in collaboration with FabD, FabG, FabZ, and FabI. Finally, the full-length fatty acid is released from the corresponding fatty acyl-ACP via hydrolysis by C16-specific thioesterase,TesA.
FabG, FabI, FabZ play significant roles in the elongation of carbon chain of fatty acyl-ACP. It is TesA that processes fatty acyl-ACP into valuable products. Therefore, we recruited these four enzymes which together would finally make it possible to efficiently produce fatty acids with longer carbon chains which are much more preferable biofuel precursors.
We have standardized the fatty acid biosynthesis related genes, TesA, FabG, FabI, FabZ, into BioBrick Parts [http://partsregistry.org/Part:BBa_K771301 Part:BBa_K771301], [http://partsregistry.org/Part:BBa_K771302 Part:BBa_K771302], [http://partsregistry.org/Part:BBa_K771303 Part:BBa_K771303], [http://partsregistry.org/Part:BBa_K771304 Part:BBa_K771304], [http://partsregistry.org/Part:BBa_K771305 Part:BBa_K771305], respectively.
Alkane biosynthetic pathway
2011 iGEM team of Washington University has introduced two enzymes that could convert ACP-Fatty Acid into alkane. The first enzyme is Acyl-ACP Reductase (AAR) which reduces long chain length acyl-ACPs into the corresponding fatty aldehydes. Acyl-ACPs are essential intermediates in fatty acid biosynthesis in every known organism, meaning that this system can work in a wide range of organisms. This long chain fatty acid acts as a substrate for Aldehyde Decarbonylase (ADC), the enzyme that removes the carbonyl group (C=O) from the fatty aldehyde, yielding an alkane one carbon shorter than the original Acyl-ACP and a molecule of formate. Since the vast majority of the fatty acyl-ACPs produced by E. coli have an even chain length, this system produces detectable amounts of only odd chain length alkanes. This study reported production of the C13, C15, and C17 alkanes, as well as the C17 alkene (unsaturated hydrocarbon). This chain length range falls well within the range of those found in diesel, so this system is theoretically able to make the alkane portion of a fuel compatible with current engines and infrastructure
File:12SJTU Alkane Biosynthetic Pathway.jpg Fig.2 shows alkane synthetic pathway that originates from cyanobacteria species. Alkane Biosynthetic pathway is combined with Fatty Acid carbon-chain elongation pathway in E.coli Design of Experiment
Mechanism beneath our idea suggests the Membrane Magic project enjoys two fundamental privileges: the Priority to Exportation and the Refinement of Interaction.
The Priority to Exportation
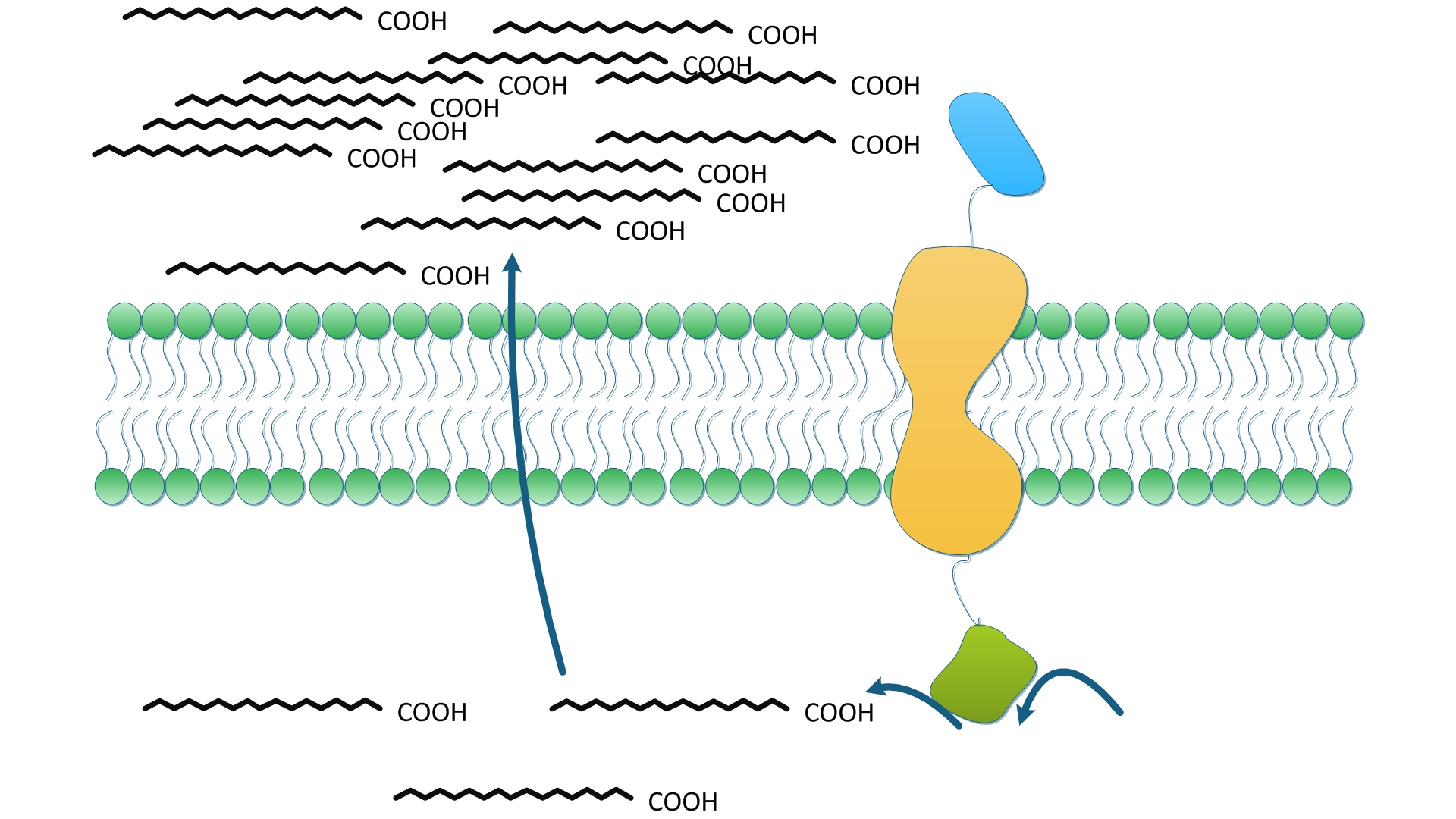 Fig.2 shows how one solo enzyme anchored onto membrane could serve to facilitate the reaction in term of the priority to exportation. The fatty acid synthesis is terminated by the hydrolysis of fatty acyl-ACP and the release of free fatty acids into cytoplasm. We suppose TesA anchored on membrane could effectively increase the concentration of fatty acids near membrane, which in turn, facilitates the transmembrane transportation of fatty acids. Higher level of fatty acids in the culture medium make it easier to obtain and purify product and more suitable for Industrialized production. On the other hand, TesA removes fatty acyl-ACP from the reaction and thus, according to Le Châtlier principle, shifts the chemical equilibrium to accelerate and to accumulate more fatty acids.
To identify and to evaluate the priority of products to exportation, controlled experiments were designed and conducted. Wild type E.coli and E.coli overexpressing diffusing cytoplasmic TesA were used as control groups. TesA was fused with Membrane Anchor 1 was expressed in experimental group.
The Refinement of Interaction
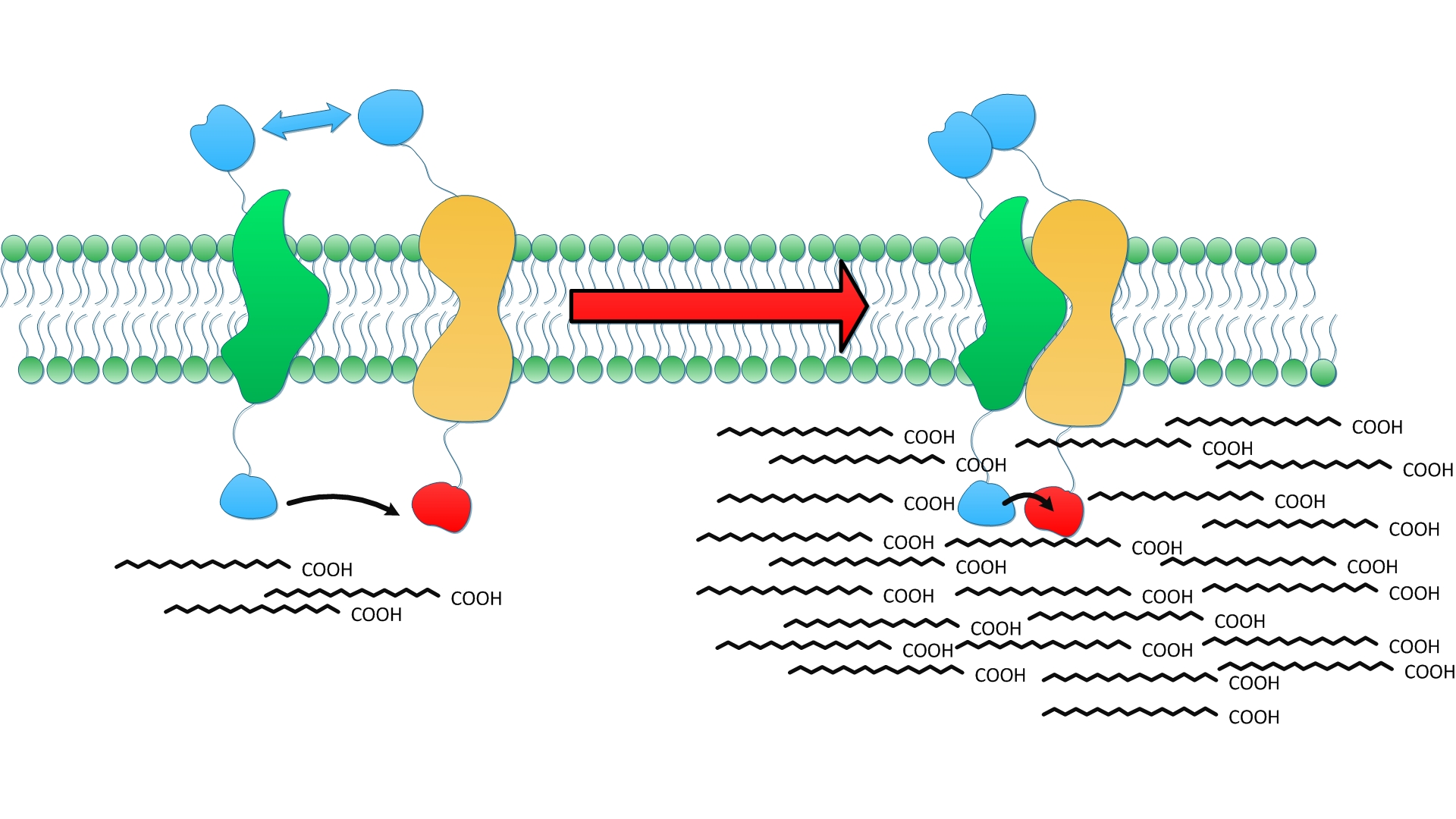 Fig.3 shows how membrane could serve to refine and stabilize the interaction upon which enzymes aggregate and cooperate. Enzymes fused with membrane anchors will be directed to the membrane as expected right after their translation. Thus, the distribution of enzymes is restricted to the 2D membrane rather than diffusing randomly throughout the cytoplasm. Due to spatial restriction, each membrane anchor is more easily to interact with each other compared to diffusing scaffold. Moreover, the interaction could be stabilized by phospholipid around transmembrane domain. We believe a tandem of enzymes involved in sequential reactions could be organized on the membrane swiftly and orderly.
To testify and to assess the refinement of interaction, controlled experiments were designed and conducted. Four enzymes are selected based on previous study. We express thioesterase(TesA) and full complement of reductive enzymes(FabG, FabZ and FabI) because previous studies showed moderate overexpression of TesA in gave rise to elevated fatty acid productivity and reductive enzymes could lead to 50% increase in fatty acid turnover. Wild type E.coli and E.coli overexpressing diffusing TesA, FabG, FabZ and FabI were used as control groups. TesA, FabG, FabZ and FabI were fused with orderly organized Membrane Anchors, aligning with corresponding reactions occurring in sequence. Four fusion proteins were expressed together in experimental group.
 Fig.4 demonstrates the construction details of Membrane Accelerator for use of fatty acid synthesis . The Parts Registered as Membrane Anchor 2 without MS2-TesA, Membrane Anchor 3-FabG, Membrane Anchor 4-FabZ, Membrane Anchor 5 without VVD-FabI are [http://partsregistry.org/Part:BBa_K771305 Part:BBa_K771305], [http://partsregistry.org/Part:BBa_K771306 Part:BBa_K771306], [http://partsregistry.org/Part:BBa_K771307 Part:BBa_K771307], [http://partsregistry.org/Part:BBa_K771308 Part:BBa_K771308], respectively.
Result and Discussion
Overview
Excitingly, the fatty acid biosynthesis was accelerated sharply by recruiting membrane accelerator. The introduction of TesA on membrane alone results in 50% increase in fatty acids yield. By gathering FabG, FabI, FabZ and TesA on the membrane, the production of fatty acid was enhanced significantly by 9 fold compared with E.coli overexpressing the same amount of corresponding cytoplasmic enzymes. The statistical data strongly support the superiority of Membrane Accelerator.
To demonstrate the priority to exportation and the refinement of interaction, two sets of controlled experiments have been set as following:
WT stands for E.coli transformed with corresponding plasmid(s) without exogenous gene.
F- stands for E.coli overexpressing corresponding free cytoplasmic enzyme(s)
M- stands for E.coli expressing corresponding enzyme(s) linked to membrane anchor(s)
Statistical data are shown as below and GC-MS data are attached to https://2012.igem.org/Team:SJTU-BioX-Shanghai/Parts
Assay of the Priority to Exportation
TesA is responsible for the hydrolysis of fatty acyl-ACP, with special affinity to C16- and C18- precursor. As Fig.5 A indicates, 20 hours after induction, E.coli with membrane anchored TesA experienced a 50% increase in C-16, C-18 fatty acids content and total fatty acids (Fig.5 B) in supernatant, compared with E.coli with free TesA.
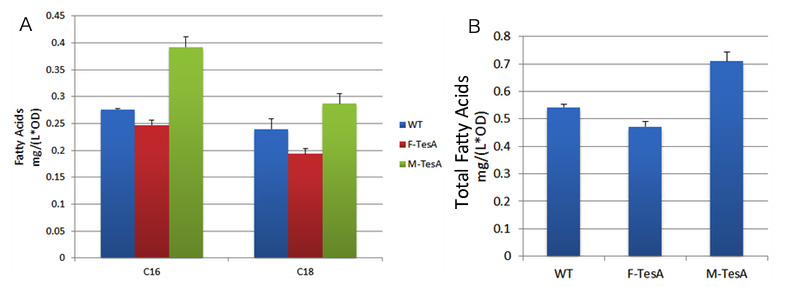 Fig.5 shows fatty acid content in supernatant of three groups to evaluate the advantages of membrane anchored TesA. A indicates C16/C18 fatty acids content among three groups. B stands for the total amount of fatty acids among three groups. Fatty acids yielded from supernatant went up to 0.71mg/(L·OD). The result supports that membrane anchored TesA could efficiently transfer fatty acyl-ACP into desirable fatty acids right beneath inner membrane, thus making it much easier for the final products to diffuse into periplasmic space.
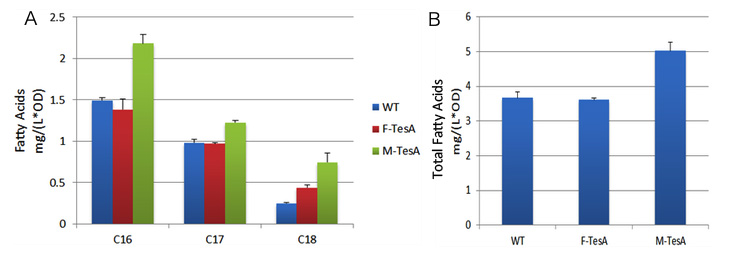 Fig.6 shows fatty acid content in the sediment of three groups to evaluate the advantages of membrane anchored TesA. A indicates C16/C18 fatty acids content among three groups. B stands for the total amount of fatty acids among three groups. Fig.6, on the other hand, shows fatty acids content in sedimentation also went up by 40%, up to 5.02mg/(L·OD). The result is still within our expectation since fatty acyl-ACP has been removed from the reaction system to form fatty acids. As a result, the chemical equilibrium shifts and more fatty acids would be accumulated.
We also witnessed a slight decrease in E.coli expressing free TesA compared with the wildtype, which testifies the statement in a previous study that high levels of TesA inhibits fatty acids synthesis activity but could enhance the activity at low concentrations.
Assay of the Refinement of Interaction
FabG, FabI, FabZ play significant roles in the elongation of carbon chain of fatty acyl-ACP. We tried to decrease the distance that intermediates need to travel by linking enzymes to aggregated membrane proteins. By this way, we accelerated the reaction and increased the turnover per unit time and finally make it possible to efficiently produce fatty acids with longer carbon chains which are much more preferable biofuel precursors.
To optimize the productivity of the system we established, we tried to combine these two privileges together, gathering TesA, FabG, FabI and FabZ through protein interaction. Notable increase in both diversity and amount of fatty acids were detected, which lends strong support to our hypothesis.
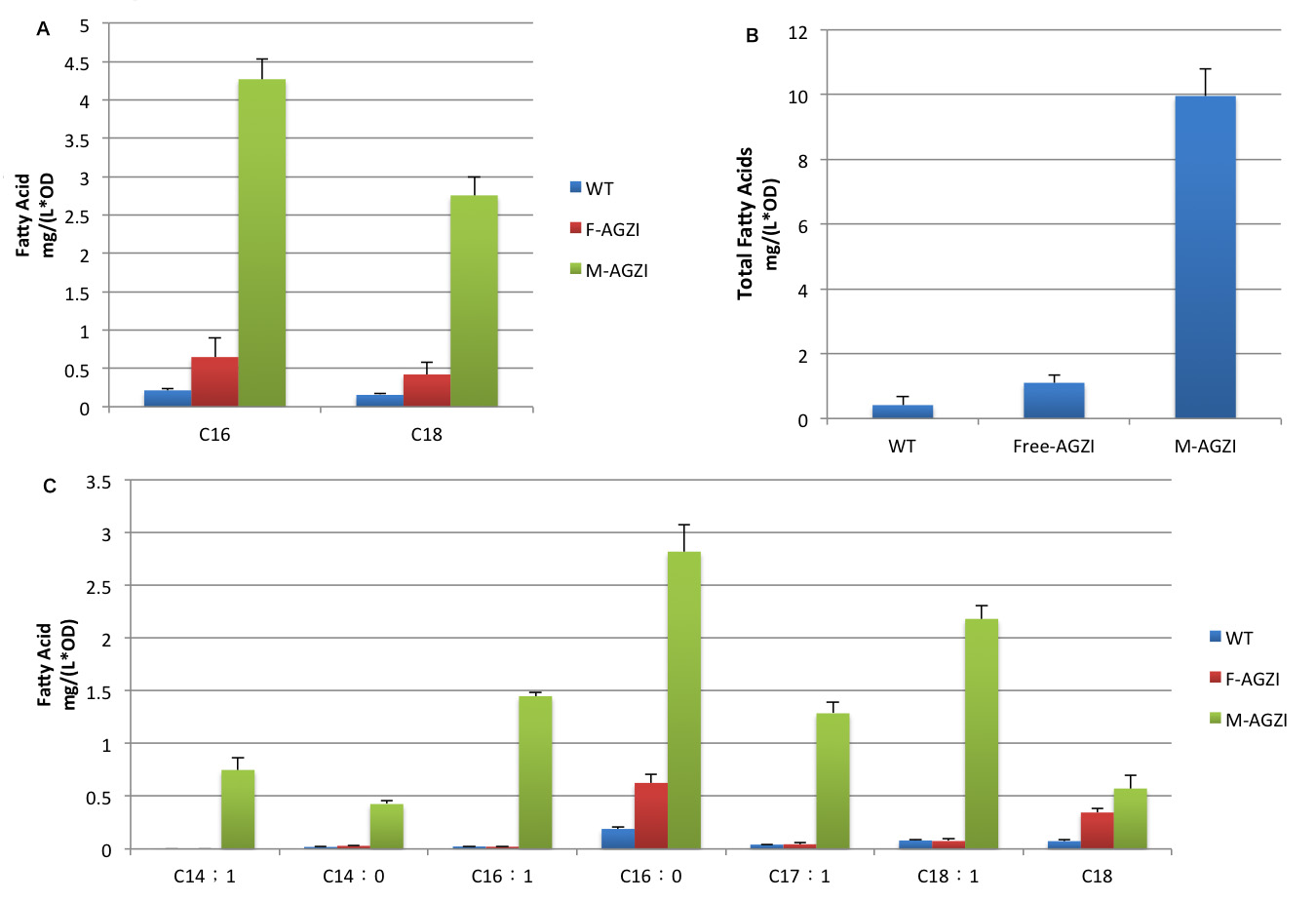 Fig.7 shows fatty acid content in supernatant of three groups to evaluate the advantages of membrane anchored TesA,FabG,FabZ and FabI. A indicates C16/C18 fatty acids content. B stands for the total amount of fatty acids. C shows the changes in products diversity. Cluster of FabG, FabI and FabZ provides C16- and C18 specific TesA with sufficient amount of fatty acyl-ACP to hydrolyze and release. Therefore, we witnessed a tremendous growth in the turnover of fatty acids with C16 and C18 skeleton in the supernatant.(Fig.7 A)
Moreover, fatty acids with C14 skeleton was first detected in E.coli expressing membrane anchored enzymes compared with ones with free enzymes. It is probably because the high productivity of membrane anchored TesA quickly released free fatty acid. Monounsaturated fatty acids also emerge in considerable amount for the first time since TesA is located so closely to the cluster of FabG, FabI and FabZ that it catches intermediate with C16 and C18 skeleton even before they are reduced.(Fig.7 C)
Augment in both diversity and amount of fatty acids led to 24 fold increase in total fatty acid yield compared with wild type and 9 fold compared with E.coli expressing diffusing cytoplasmic enzymes. (Fig.7 B) The exciting result convincingly proves that membrane scaffold enhances protein interaction and cluster of enzymes makes reaction faster to a surprising extent. Products accumulated near inner membrane can traval a shorter distance to be exported to membrane.
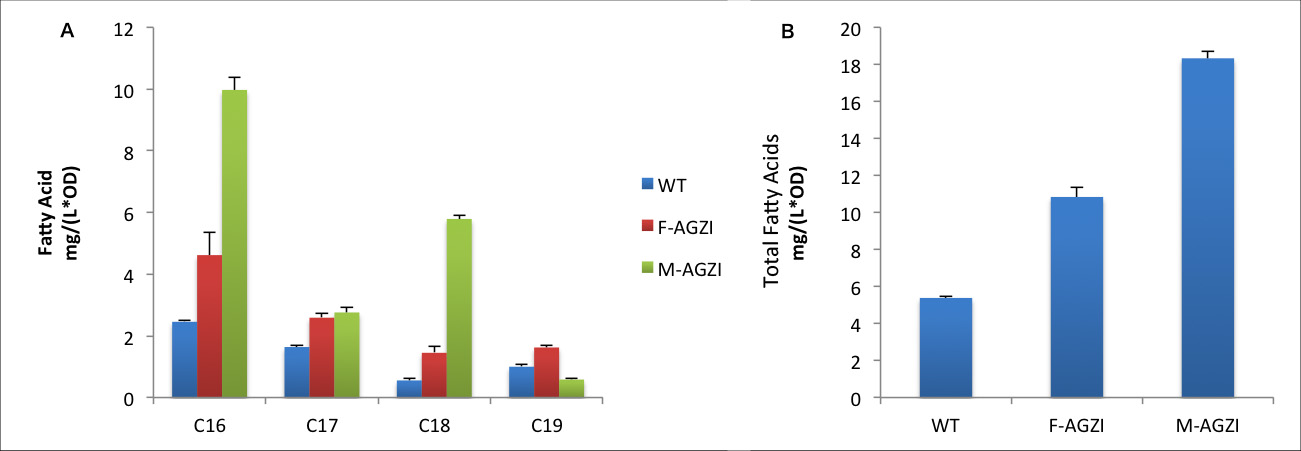 Fig.8 shows fatty acid content in sediment of three groups to evaluate the advantages of Membrane Accelerator. A indicates C16/C18 fatty acids content. B stands for the total amount of fatty acids. The fatty acids yields increase in sediment is relevantly moderate. The reason might be that membrane anchored enzymes facilitate exportation of large quantity of fatty acids so the amount of fatty acid remaining in cytoplasm is limited. It is notable that C19 fatty acids only exist in cytoplasm because large molecular weight of C19 fatty acid prevents it from exportation and finally they are trapped in the cell. C19 fatty acids yield in Membrane Accelerator Group is even less than Group with diffusing cytoplasmic enzymes. We suppose it is caused by the high efficiency of TesA which leads to quick release of free fatty acid.
Future Direction
The universal tool we developed is pointing at a promising application prospect in biofuel production. Further optimization and development could make it even more powerful.
Regardless of any optimizations with respect to strains, growth media, temperature etc., to take proper proportion of each enzyme into consideration before using Membrane Magic guarantees qualitative improvement of turnover. For instance, if monounsaturated fatty acids are less preferred than saturated ones, we can introduce extra enoyl reductase FabI into former cluster to help reduce monounsaturated fatty acids. Actually, we can precisely adjust the effective amount of certain enzyme in a cluster to achieve optimized proportion. Therefore the maximum yield could be significantly higher.
Arabidopsis thaliana fatty acid thioesterase A(AtFatA) is another candidate in chain termination reaction. AtFatA specifies in long-chain fatty acids(C18 and longer), thus providing deeper potential in biofuel production. We can even yield diesel instead of fatty acids for E.coli simply by replacing TesA with an acyl-ACP reductase(AAR) and an aldehyde decarbonylase(ADC). The cluster of FabG, FabI and FabZ can supply adequate amount of acyl-ACP, making it possible to mass produce alkanes via biosynthesis.
The priority to exportation could be further developed when fatty acid transportor is introduced right next to thioesterase. Final products can bind to transportor the moment it is released from thioesterase.
The most fascinating scenario is that we perform the complete Membrane Magic within a single cell. All the possibilities mentioned above or not, could be integrated and manipulated by Membrane Rudder.
|  "
"








{kind=link}