Biological systems function on a great variety of different integrative mechanisms which include post-transcriptional attenuation. We believe that Synthetic Biology is at its most effective when these natural mechanisms are taken and applied in novel situations. This ethos we have sought to emulate by creating our own mechanism of post-transcriptional attentuation; the Comparator Circuit.
In a previous project, the apparent lack of specificity of the promoter BioBrick we looked to improve upon proved to be a minor difficulty that we felt had not been addressed previously in the Registry. Thus we decided to tackle the issue by devising a way of quantitatively measuring the output of NO with the non-specific promoter we were using through a novel gene regulation system.
Using pairs of BioBricks that result in the complimentary binding of a pair of otherwise standard promoter and reporter constructs, a subtractive effect can be gained to result in altered translation relative to the availability of various substrates within the chassis environment.
|
Imagine you have two promoter biobricks, which both have the ability to sense the same substrates, but, critically, one of these promoter biobricks senses an extra substrate - the substrate you are interested in. The aim of the Comparator Pair of biobricks we have created is, when each of the pair is ligated to one of the promoter biobricks, to integrate these promoter outputs to result in a subtractive system that produces a quantitative output of the substrate of interest only.
|

|
|
When these promoter/comparator duplexes are added to different reporter proteins, the translation of these reporters are sequestered when both sequences are transcribed due to the presence of the same substrate.
|

|
|
So, for instance, if one of the promoters produces twice as many mRNA transcripts due to the extra substrate it is sensing...
|

|
Amy Congdon
NRP UEA iGEM Artist
Amy Congdon completed her BA in Contemporary Textile Practices, at Norwich University College of Arts, and then went on to do her masters in Textile Futures, based at Central Saint Martins in London. Amy has played an important role as the team’s artist, since she got in touch with the team a few months before they started on their project. She is fascinated between the links of design, craft and science, and has used her interest to help the team to engage with the public. The team are very thankful to have her on board for her creative input, as well as her dedication and time to the team’s project. Amy has been very active within our human outreach program; from designing the team’s eye catching logo, to directing and editing their film.
|

|
This is done via the comparator pairs design, which facilitates complete binding of both mRNAs ribosome binding sites when they are both present in the chassis' transcriptome.
The video below describes the project in further detail:
An important part of the Comparator Circuit idea is the fact that there is potential of other Comparator Pairs being made to result in a family of biobricks existing on the registry. Different Comparator Pairs could thus be expressed simultaneously in the cell, working with different promoters to quantify a number of different substrates in tandem within the same chassis.
Planning
Figure 1 - Both BioBricks of the Comparator Circuit bound together.
Assembling the various gene constructs was not without its challenges. Due to the fact that complimentary ‘zips’ within the sequence were designed to surround the ribosome binding site it was often the case that the DNA sequence would form unwanted secondary structures that could serve to inhibit the translation of the mRNA in its uncoupled state.
Therefore, when designing the DNA, care was taken to avoid these structures obstructing sequences required for translation of the mRNA. Simultaneously, only common codons for the chassis of interest, E. coli, could be used and these codon had to code for amino acids that were unlikely to change the function of the protein product. This is required since the zip sequences extend past the translational start codon, thus our construct will add a small N-terminal tag to any reporter protein it was attached to.
Using IDT's very helpful online tool, Oligo Analyser, we were able to produce sequences in both constructs required in the Comparator Circuit system that, theoretically, bound together when transcribed at around the same within the chassis, yet left the ribosome bind site open and unfolded when either strand of mRNA lacks its counterpart to complimentarily bind to. The software produced figures demonstrating the likely secondary structure of the constructs made, and their Gibbs Free Energy value at specific temperatures. From this, trial and error eventually resulted in the construction of two BioBricks that have free ribosome binding sites when in isolation, but bind to sequester translation of both mRNAs when bound together (Figure 1).
Due to the stop codon present in the scars of Assembly Standard 10 BioBricks, we decided that our constructs would have to be an Assembly Standard 23 BioBrick. Although the use of Bioscaffolds produced by previous iGEM teams was considered, time constraints meant that changing the Assembly Standard we put our BioBricks in would be the most convenient solution.
Experiments
Due to the limited time we had available, it was decided fairly early on that the comparator circuit BioBricks could only be characterised theoretically. However, our confidence in the system meant that we felt obliged to synthesise these BioBricks and submit them to the registry. It is our hope that we will be granted lab space after the regional jamboree to characterise these BioBricks and submit our findings onto their registry experience page in due course. In preparation for this, the team have devised a provisional experimentation design for these characterisation studies.
Future Experiments
Further cloning experiments hope to be carried out, ligating each comparator circuit biobrick with a different reporter and promoter. Initially, for proof of concept, the promoters used would be simple promoters that have overlapping specificity, but one substance will only act as an inducer for one of these promoters (e.g. the pYEAR and SoxR promoters, which both sense small nitrogen species, but SoxR also senses superoxides).
Reporter biobricks would also be ligated, with these reporters being as far apart in wavelength of the florescence produced as possible. Both DNA sequences, each containing one half of the comparator circuit system, with be transformed into an E. coli chassis and the amount of either fluorophore (as seen via fluorometer studies) would indicate the translation of either sequences. It is hoped that in simulated environments containing higher levels of, using the example above, superoxides, the corresponding fluorescent protein will be translated over its counterpart.
Ultimately it is hoped that we would be able to ligate our comparator circuit BioBricks to NO-sensing promoters and effector enzymes to control NO levels/reporters to detect NO levels quantitatively - which is where our fictitious brand Quanticare comes in. Should we have time, we'd like to investigate if the transfection of mammalian cells with these constructs, as seen in our science-fiction self-diagnostic tattoos, is plausible via the transfection of cancer cell lines with the DNA constructs.
Beyond the addition of promoters and transformation into a bacterial chassis, further experiments which would need to be carried out are:
1) A growth study into whether the incorporation of the comparator circuits affect the growth of the chassis compared to growth without can be tested
2) A fluorescence study into whether the comparator circuit functions as expected. This would involve the use of a fluorometer and FACs to obtain a comprehensive view of the proteins.
3) A cytotoxic study involving a cytotoxic assay similar to the one carried out on BM and MB, however, this will test whether the accumulation of silenced RNA transcripts will affect the cell.
4) Study into the sensitivity of the promoters- this be a measure of threshold levels of a promoter before it expresses.
Theoretical Characterisation
The idea of the comparator circuit is to provide a modular method of signal integration that can produce a sensor which can specifically and quantitatively measure different chemical species within the cell using non specific promoters. Through mathematical modelling, an equation has been assembled which can predict the expression of each of the reporter proteins such as RFP and CFP.
Figure: Theoretical equation to predict the degree of expression of Construct 1 and 2.
The full equation has been laid out in a way that is relevant only to Construct 1, however, the numbers can be reversed to be relevant to Construct 2. For ease of explanation, everything described will be relevant to Construct 1.
E = Proportion of expression rate of Construct 1 when both constructs are transcribed (i.e. there is knockdown of one construct) relative to the non-knocked down expression of Construct 1 when only Construct 1 is expressed.
A = The rate of transcription of Construct 1 as a proportion of the maximum transcription rate. As a proportion this is measured on a scale of 0 - 1. As an example if the rate of transcription is half of the maximum rate, rate would be 0.5 (arbituary units). It can be assumed the rate of transcription of construct 1 and 2 due to cellular components (e.g. RNA polymerase) is the same, however, the rate of transcription initiation will dictate the transcription rate. The initiation is reliant on the chemical species interacting with the transcription factor which binds to the promoter (i.e. nitric oxide,nitrates,nitrites to PyeaR). The '1' and '2' refer to the Construct 1 or 2 and hence the promoter and the measured fluorescent protein attached (e.g GFP, RFP, CFP, etc).
L = The length of the Construct 1 in the DNA form that is transcribed (i.e the leader and protein coding region).
Note: Leader refers to the section of RNA at the start of the mRNA that is not translated but has an effect on translation rate.
C = The rate of transcription. Assuming the rate of transcription of Construct 1 and 2 are the same because the same ribosomes and RBS are involved.
T = Half life of Construct 1 when only Construct 1 is present; the natural half life of Construct 1.
K = A constant of the biological system. This can only be measured through observation.
The full equation is modelled on the basic equation of:
where E is the rate of expression and E(A1) is the same as that explained above.
The additional complexity factors in less assumptions, and mimics a biological system, more closely. Below is a breakdown of the full equation.
This refers to the number of Construct 1 RNA transcripts undergoing transcription at any one time. The length of DNA is particularly important when the chassis is bacterial. In bacteria, as there is no true nucleus, translation occurs simultaneously with transcription. Transcription affects the probability of interaction between construct 1 and 2 and therefore, they are less likely to be translated. As the measurement of fluorescence is the output directly related to the rate of translation, the overall equation measures translation, however, translation rate iis dependent on rate of transcription and degree of knockdown, and hence transcription is factored in here. L/C is the period of time taken for transcription to take place. It is the time in which translation can be initiated but it is unlikely that the two leaders will bind to one another
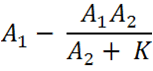
This part of the equation is the deduction of the knockdown of Construct 1 when there is Construct 2 expression and interaction. The biological constant, k, factors in that not all of construct 2 that is expressed will interact with construct 1 and vice versa. Hence, both exist despite construct 2 existing in small quantities. We believe that depending upon the assembly of the orientation of the two constructs within the plasmid, the interaction and hence the binding efficiency can be altered vastly. If the genes have opposite orientations, so that the termination sites are very close then the reduction of distance will increase the chances of interaction and hence make the sensory system more accurate.
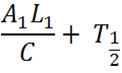
This part of the equation encompasses the natural half life of Construct 1 when it alone is expressed (i.e. no expression of or interaction with Construct 2). As described before in the modelling from the basic equation, this is the lower part of the equation and puts it in perspective of Construct 1 and gives expression as a porportion of the maximum transcription. The half life is also Construct 1's half life.
So to bring it all together; the top half of the equation indicates the degree of translation of the RNA transcribed by the first promoter under any particular transcription rate of the two promoters in arbitrary units. To make this into a meaningful output it is divided by the maximum translation rate at that rate of transcription to equal E(A1); this indicates the degree of attenuation of one RNA from the other.
Future Applications
The Comparator Circuit has the potential to have real world applications, particularly, as QuantiCare shows, in medicine. To give an example, by monitoring blood sugar levels quantitatively via specific promoters, the sequence following the comparator circuit could encode for insulin. This could be transfected into human cells and could be used to alleviate the symptoms associated with type I diabetes.
Moving back to the specific nitrogen sensor, attaching these promoters to the comparator circuit biobricks and a gene for the synthesis of nitrogen reductase could result in a positive feedback loop to result in the tumour reducing in size. Macrophages naturally sense the presence of tumours in the body via their emission of nitric oxide. This could be taken one step further by adding nitrogen reductase to this system, where an excess of nitric oxide in the tumour environment. NO was seen to have cytotoxic properties in large amounts and, thus, a positive feedback loop could result in tumour apoptosis.
 "
"







